Динамічні показники популяції
До динамічних показників популяції належать типи росту і типи динаміки чисельності (чи щільності) популяції.
У загальному вигляді рівняння динаміки чисельності популяції має вигляд:
швидкість зміни чисельності = (народжуваність + швидкість іміграції) — (смертність +швидкість еміграції).
До основних динамічних показників популяції належать: швидкість народжуваності (чи просто народжуваність) - кількість особин, що народжуються в популяції (∆N) за певний проміжок часу ∆t; для оцінки народжуваності та порівняння за цим показником різних популяцій зручно користуватися питомою народжуваністю ∆N∕∆t. Смертність - величина, протилежна народжуваності.
5.4.1. Типи росту популяцій. J- подібний та S- подібний ріст
Розрізняють два основні типи росту чисельності (чи щільності) популяцій - J-подібний і S-подібний, названі так за характером кривих росту (рис. 6.1).
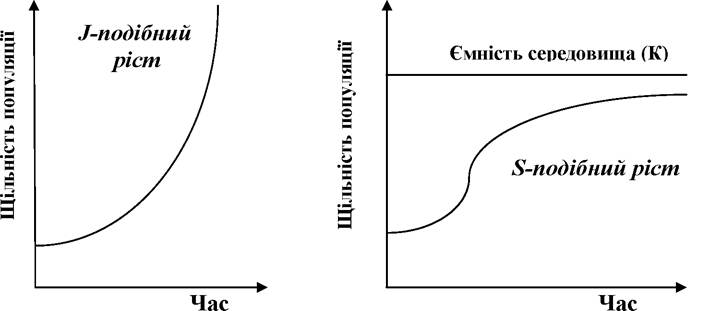
Рис. 5.6. Типи росту популяцій
Якщо чисельність особин зростає з відносно постійною швидкістю, то має місце типовий J- подібний ріст.
При цьому миттєва швидкість росту визначається таким чином:
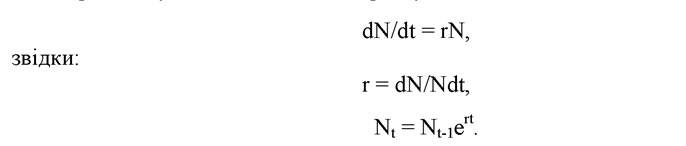
Якщо r = const, то ріст відбувається за експоненційним законом.
Якщо величини чисельності наводяться в логарифмічному масштабі, то графік експоненційного росту набуває вигляду прямої лінії. Тому інколи експоненційний ріст називають логарифмічним.
Рівняння експоненційного росту в логарифмічній формі має вигляд: 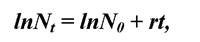 тобто є рівнянням прямої, а коефіцієнт r характеризує кут нахилу її до осей.
тобто є рівнянням прямої, а коефіцієнт r характеризує кут нахилу її до осей.
Біологічний сенс коефіцієнта r (від англ. rate - рівень) досить цікавий.
По суті, це біотичний потенціал популяції - миттєва швидкість зміни чисельності чи щільності популяції.В кожний конкретний момент часу його можна розглядати як різницю між миттєвою народжуваністю та миттєвою смертністю:
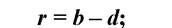
де: r - біотичний потенціал популяції; b - рівень народжуваності (birth-rate); d - рівень смертності (death-rate).
Таким чином, у кожний момент часу популяція може змінювати свій біотичний потенціал або шляхом зміни рівня народжуваності, або рівня смертності, або ж обох цих показників одночасно.
S- подібний ріст популяції
Для опису цього типу росту можна використовувати багато рівнянь, але найбільш популярним виявилось найпростіше з них - так зване логіс- тичне рівняння. Вперше його запропонував бельгійський математик П.Ф. Ферхюльст для опису росту народонаселення (Verhulst, 1838). Цим рівнянням користувалося багато дослідників, проте з часом про нього забули. Потім його «перевідкрили» американські дослідники Пірл та Рід (Pearl, Reed, 1920), які, втім, через рік визнали пріоритет Ферхюльста.
В основі логістичної моделі лежить припущення про лінійне зниження швидкості питомого росту ra = dN/Ndt зі зростанням чисельності N, причому ця швидкість стає рівною нулеві при досягненні певної (граничної для даного середовища) чисельності - К. Тобто якщо N = K, то ra = 0.
Логістичне рівняння в диференціальному вигляді:
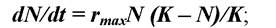
де: rmax - константа експоненційного росту, який міг би спостерігатися в початковий момент зростання чисельності; К - верхня асимптота S'- подібного росту. Біологічний сенс її можна ототожнити з поняттям “ємності середовища” для популяції. Ємність середовища (для популяції) — це максимальна щільність популяції, яка може підтримуватися ресурсами даної екосистеми.
В інтегральній формі погістичне рівняння має вигляд:
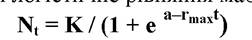
де: Nt - чисельність популяції в момент часу t, e - основа натурального логарифму, К - верхня асимптота S - подібного росту, або ємність середовища.
Варто звернути увагу, що в рівнянні S-подібного росту значення біотичного потенціалу популяції (r) постійно змінюється, прямуючи до нуля, в той час як при J-подібному рості значення r постійне в будь-який момент часу; тому в рівнянні S-подібного росту використовують не r, а rmaxЗауважимо, що термін “логістична крива” був запропонований П.-Ф. Ферхюльстом без будь-яких пояснень. У французькій мові того часу слово “logistique” означало “мистецтво обчислення”.
Знання особливостей росту певної популяції конче необхідні для її раціональної експлуатації. Зокрема, за умови S-подібного росту, максимальну продукцію можна отримувати при підтриманні щільності популяції на рівні, близькому до точки зміну радіусу кривизни S-подібної кривої (рис. 5.6), тобто за найвищої швидкості росту даної популяції. Цей же підхід дозволяє визначити верхню межу вилучення особин з популяції (що в свою чергу залежить від задач оптимізації використання певної популяції).
5.4.2. Типи динаміки чисельності популяцій
У загальному вигляді співвідношення процесів, що визначають динаміку чисельності популяції, можна записати таким чином:
Зміна чисельності популяції = (народження особин + імміграція) — (загибель особин + еміграція).
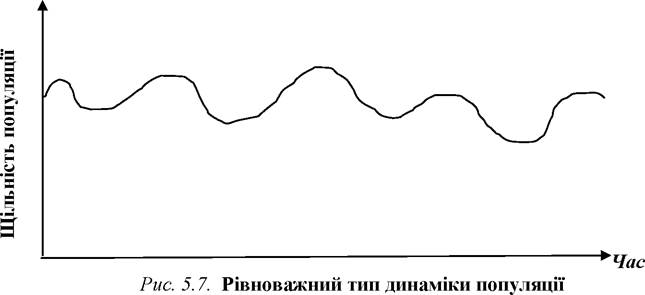
За особливостями динаміки чисельності всі популяції можна поділити на дві основні групи: рівноважні та опортуністичні. Перші - це популяції, щільність яких не має різких перепадів, вона зазвичай коливається в певних межах навколо якогось середнього значення. Зазвичай рівноважний тип динаміки щільності притаманний популяціям тих видів, представники яких відзначаються відносно великими розмірами, значною тривалістю життя, тривалим життєвим циклом (рис. 5.7).
Щодо особливостей росту чисельності (щільності) рівноважних популяцій, то, зазвичай, їм притаманний S- подібний ріст, тоді як опортуністичним - J-подібний (що і дає їм можливість вибухоподібно збільшувати свою щільність за відповідних умов).
Опортуністичні ж популяції характеризуються різкими коливаннями щільності.
Такий тип динаміки характерний багатьом видам, яких називають «шкідниками» різних господарств тощо, зокрема загальновідомі спалахи щільності сарани, мишовидних гризунів тощо. Графіки динаміки їх чисельності (чи щільності) часто нагадують електрокардіограму чи енцефалограму (рис. 5.8).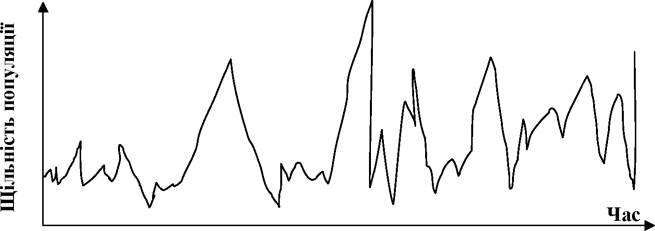
Рис. 5.8. Опортуністичний тип динаміки популяції
Щодо причин коливання щільності популяцій існує безліч теорій. Зокрема, навіть за стаціонарних умов у популяції буде спостерігатися певна цикліка її щільності, пов’язана з періодичністю розмноження, тривалістю життя тощо (рис. 5.9)
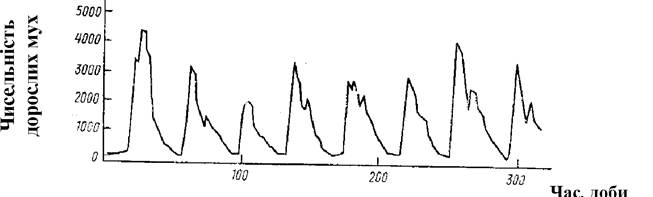
Рис. 5.9. Циклічні коливання чисельності мух Lucilia cuprina в лабораторній популяції, що культивується за постійної кількості корму (за Nicholson, 1954)
Часто причини коливань чисельності чи щільності популяцій пояснюють різними зовнішніми (для даної популяції) впливами. Досить популярною є теорія коливань, що ґрунтується на взаємовпливах хижака і жертви. Зокрема, майже в усіх підручниках з екології наводять приклад коливання чисельності зайця і рисі в Канаді (за матеріалами заготівель шкір компанією Гудзон-Бей (MacArthur, Connel, 1966) (рис. 5.10).
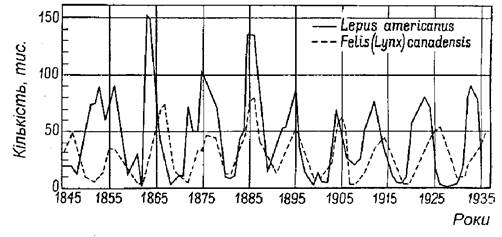
Рис. 5.10. Динаміка чисельності зайця Lepus americanus і рисі Lynx canadensis у Канаді (за матеріалами заготівель шкір компанією Гудзон-Бей)
Проте у більшості випадків встановити, що є причиною, а що - наслідком - досить важко. Зокрема, динаміку чисельності жертви можна пояснити тиском хижаків, проте з такою ж вірогідністю можна стверджувати, що динаміка популяції хижака (рис. 5.10) лише відповідає на зміну чисельності жертви, причина якої не відома.
І лише в кількох дослідженнях можна з високою вірогідністю говорити про причини та наслідки динаміки щільності популяцій.
Досліджуючи причини динаміки щільності масових видів планктонних ракоподібних на водосховищі в нижній течії р. Замбезі в тропічній Африці, польський гідробіолог Глівіч виявив дивний зв’язок між фазами місяця і динамікою щільності масових видів планктонних ракоподібних (Bosmina Iongirostris, Diaphanosoma excisum, Eudiaptomus sp., Mesocyclops leuckarti) (Gliwicz, 1986). Зокрема, було встановлено, що чисельність цих ракоподібних протягом року закономірно коливається, причому максимуми щільності завжди припадали або ж на повнолуння, або ж за кілька діб до нього. Глівіч з’ясував причину цього явища: інтенсивне виїдання ракоподібних уночі за повного місяця невеликими (до 8 см завдовжки) прісноводними сардинами лімнотрисами (Limnothrissa miodon). Планктонні ракоподібні в цій водоймі, як і в багатьох інших, здійснюють добові вертикальні міграції, піднімаючись в темну частину доби до поверхні - в шари, багаті їжею, а в світлу частину доби залишаються у придонних шарах на значній глибині, де через незначний рівень освітленості у них значно менший ризик стати жертвою риб. Риби також здійснюють вертикальні міграції - скопичення їх пливуть слідом за максимальним скопиченням планктону (що було встановлено з використанням ехолотного дослідження). У безмісячні ночі ско- пичення риб розсіюються, живлення не спостерігається. З наближенням до повного місяця скопичення риб починають зберігатися й на ніч, а максимальна інтенсивність виїдання планктону спостерігається за максимального освітлення води - повний місяць та кілька діб до і перед ним (рис. 5.11).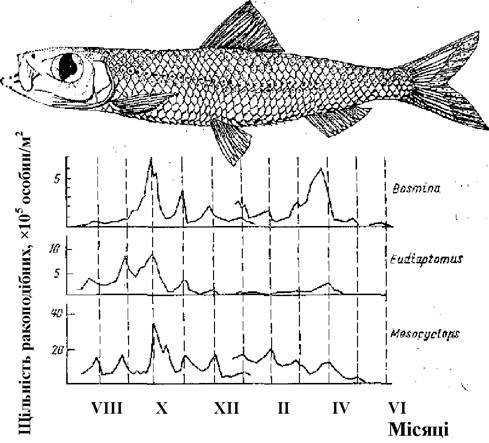
Рис. 5.11. Спряжені з місячним циклом коливання щільності трьох видів планктонних ракоподібних у водосховищі Кагора-Басса на р. Замбезі. Вгорі- Limnothrissa miodon, яка активно виїдає зоопланктон у місячні ночі (пунктирними вертикальними лініями відмічені доби повного місяця)
Знання закономірностей динаміки щільності популяцій конче необхідне для розробки квот виловів риб певних видів, ліцензування відстрілу окремих видів тварин, лісокористування, прогнозування спалахів чисельності видів, які є переносниками найрізноманітніших захворювань, «шкідниками» лісового і рибного господарств тощо.
Популяційна екологія досягла значного розвитку значною мірою саме завдяки її практичній спрямованості, адже раціональне використання популяцій, як складова раціонального збалансованого природокористування, має опиратися саме на наукові дослідження особливостей їх відтворення у конкретних екосистемах. Адже, як підкреслював О.В. Яблоков, «визначення меж відведення особин та біомаси популяцій є центральним завданням природокористування. Популяція є першим акцептором, який сприймає всю різноманітність порушень, що вносить в навколишнє середовище діяльність людини. Якщо антропогенна діяльність навіть і не направлена прямо на ту чи іншу популяцію, вона опосередковано сприймає такі дії. Перевищення порогів використання популяцій завжди загрожує серйозними наслідками».
Тому раціональне використання кожної популяції має ґрунтуватися на детальних відомостях щодо її особливостей, зокрема продукційних та циклічних характеристиках.
5.4.
Еще по теме Динамічні показники популяції:
- Статичні показники популяції
- Показники економічної і соціальної ефективності природоохоронних заходів
- 4. ПРОБЛЕМА ДЖЕРЕЛ СОЦІАЛЬНОЇ ДИНАМІКИ. КОНСТРУКТИВНІ ТА ДИНАМІЧНІ СИЛИ НАУКОВО- ТЕХНІЧНОГО ПРОГРЕСУ
- Кількісна оцінка якості середовища за продукційно- енергетичними показниками біосистем
- Популяції. Структура та динаміка популяцій
- ЧИСЕЛЬНІСТЬ І ЩІЛЬНІСТЬ ПОПУЛЯЦІЇ
- ЕКСПЛУАТАЦІЯ ПОПУЛЯЦІЇ
- ПРОДУКТИВНІСТЬ ПОПУЛЯЦІЇ
- Популяції: означення та параметри
- СТАТЕВА І ВІКОВА СТРУКТУРИ ПОПУЛЯЦІЇ
- ПОНЯТТЯ ЕКОЛОГІЧНОЇ СТРУКТУРИ ПОПУЛЯЦІЇ
- ПРОСТОРОВА СТРУКТУРА ПОПУЛЯЦІЇ
- ХАРАКТЕР І РОЗМІЩЕННЯ ОРГАНІЗМІВ У ПОПУЛЯЦІЇ
- Поняття популяції
- Вплив забруднення довкілля на популяції та екосистеми