Речовинна структура екосистем. Біогеохімічні колообіги
Між біотичними і абіотичними компонентами екосистем відбувається безперервний обмін різноманітними хімічними елементами, обумовлений взаємодією біологічних і геохімічних процесів.
На відміну від потоку енергії, колообіг речовин в екосистемі відносно замкнений, а для біосфери в цілому біогеохімічні колообіги можна розглядати як практично замкнені.Саме колообіг речовин є найважливішим механізмом забезпечення цілісності будь-якої екосистеми, подібно тому, як кровообіг забезпечує цілісність організму тварин. Проте, на відміну від біосфери, колообіги хімічних елементів окремих екосистем пов’язані з іншими екосистемами. Саме за ступенем замкненості біогеохімічних колообігів можна характеризувати ступінь своєрідності, чи замкненості (ізольованості) тієї чи іншої екосистеми.
Близько 40 хімічних елементів, що входять до складу живої речовини, постійно циркулюють між живим і неживим компонентами екосистеми.
Загальна формула “живоїречовини ”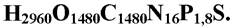
У порівнянні з неживим компонентом жива речовина більш карбок- сильована, гідрогенізована і гідратована.
Біогеохімічний колообіг (БГХК) - це шлях циркуляції хімічного елемента по відносно замкненому ланцюжку, ланками якого є живі й неживі компоненти.
У БГХК розрізняють активний фонд елемента - та його частка, що перебуває у складі живої речовини або у стані руху від (чи до) неї та резервний фонд (депо) - відносно нерухомі запаси даного елемента. Між активним та резервним фондом постійно здійснюються певні потоки, які забезпечують підтримання рівноваги в системі в цілому.
За ознакою головного депо БГХК поділяються на дві основні групи: колообіг газоподібних речовин (КГР) та осадовий цикл (ОЦ).
Колообіг газоподібних речовин включає всі елементи, які бодай на одному з етапів утворюють газоподібні сполуки.
Елементи осадового ж циклу до складу газів не входять. І в цьому між ними пролягає суттєва відмінність. Резервний фонд елементів КГР знаходиться в атмосфері та гідросфері (в розчиненому стані), тому їх колообіг, як правило, добре збалансований.Щодо елементів ОЦ - то їх колообіг власне і колообігом назвати не зовсім коректно - він розбалансований і однонаправлений, тобто елементи вилуговуються з гірських порід, входять до складу живих організмів, виходять з них, але врешті-решт в тій чи іншій формі вони змиваються з водозбірного басейну і виносяться водотоками в озера, моря, океани, де і захоронюються в донних відкладах. Зворотній винос цих елементів пов’язаний, в основному, з видобуванням морських продуктів, але це у більшості випадків складає лише відсотки чи їх частки від загальної величини виносу елемента. І лише в масштабах геологічного часу - коли дно моря стане суходолом - цей тип руху елемента можна назвати колообігом.
8.2.1. Колообіг газоподібних речовин
Прикладами колообігу газоподібних речовин є БГХЦ нітрогену, карбогену, оксигену, сірки та інших елементів, які утворюють газоподібні сполуки.
8.2.1.1. Колообіг нітрогену
Азот складає 78% атмосфери, але переважна більшість живих істот не здатна безпосередньо використовувати цей запас азоту. Включення азоту в сполуки, які використовуються організмами, називається його фіксацією.
Цикл азоту складається з чотирьох основних етапів:
1. Азотфіксація.
2. Нітрифікація.
3. Амоніфікація.
4. Денітрифікація.
Азотфіксація - процес зв 'язування молекулярного азоту в азотисті сполуки. Вона буває абіогенною і біогенною. Перша здійснюється внаслідок розрядів блискавок і фотохімічних процесів.
На початок 50-х років вважалося, що азотфіксацію здійснюють лише кілька груп поширених у природі мікроорганізмів - вільноживучі бактерії Azotobacter (аероб), Clostridium (анаероб); симбіотичні бульбочкові бактерії бобових Rhizobium; ціанобактерії - Anabaena, Nostoc.
Потім азотфіксацію виявили у пурпурових бактерій Rhodospirillum та інших представників фотосинтезуючих бактерій.
Встановлено, що актиноміцети в кореневих клубнях вільхи і деяких інших рослин фіксують азот неменш ефективно, ніж бульбочкові бактерії у бобових рослин. Із кожним роком список азотфіксаторів розширюється.
Азотфіксацію здійснюють авто- і гетеротрофні організми як в аеробних, так і в анаеробних умовах.
Серед анаеробів численні види р. Clostridium, деякі метаноутворюючі, сульфатредукуючі й фотосинтезуючі бактерії (Горленко и др., 1977).
Із аеробних форм найбільше значення мають бактерії родини Azotobacteriaceae (Azotobacter та ін.) і особливо ціанобактерії (Anabaena, Aphanizomenon, Nostoc, Oscillatoria, Microcystis, Nodularia, Gleocapsa та ін.). Здатність до азотфіксації притаманна деяким воднеокислюючим і фо- тосинтезуючим бактеріям р. Chlorobium, Chromatium та ін. Для активації азоту автотрофи використовують енергію фото - і хемосинтезу, гетеротро- фи - енергію, яка міститься в органічній речовині, що ними споживається.
За даними Делвіч (1965, 1970) в біосфері фіксація азоту з повітря складає на суходолі в середньому понад 1 г/м2 за рік. За даними Фогга (1955) в родючих областях біологічна фіксація азоту досягає 20 г/м2 за рік. В основному - це біологічна фіксація, і лише незначна частина (у помірних широтах не більше ніж 35 мг/м2 на рік) фіксується внаслідок електричних розрядів і фотохімічних процесів.
У Балтійському морі і Рибінському водосховищі інтенсивність азотфіксації досягає 0,8-1 г/м2 на рік (Саралов, 1979). Всього в гідросфері щорічно фіксується близько 10 млн т азоту.
Перший етап азотфіксації - активація азоту: розщеплення його на два атоми, на що витрачається 672 кДж/моль. Наступна фаза реакції відбувається за схемою: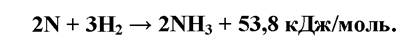
Реакція каталізується ферментом нітрогеназою, яка руйнується за наявності кисню, що вельми суттєво для екології азотфіксаторів. Так, у ціанобактерій азотфіксація здійснюється в гетероцистах, потужна оболонка яких ізолює їх вміст від доступу кисню.
Висока нітрогеназна активність виявлена у вмісті кишківника морських їжаків, особливо за умов, коли вони не отримували азот з їжею: азот, що фіксувався мікроорганізмами, включався в клітини морських їжаків (Guerinot, Patriquin, 1981).Наявність сполук нітрогену гальмує азотфіксацію, оскільки мікроорганізмам енергетично вигідніше використовувати готові речовини, ніж синтезувати їх. Фіксація азоту потребує значних витрат енергії, оскільки багато її йде на розривання потрійного зв’язку в молекулі азоту. Бактерії в бульбочках бобових витрачають на біофіксацію 1 г азоту близько 10 г глюкози. Слід зауважити, що і виробництво азотних добрив є найбільш енергомістким з усіх виробництв мінеральних добрив.
Нітрифікація полягає в окисненні аміаку до азотистої кислоти, а потім окиснення нітритів, до нітратів,:
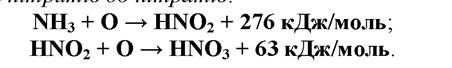
135
Перша реакція здійснюється бактеріями роду Nitrosomonas, друга - роду Nitrobacter. Обидва роди бактерій використовують вивільнену енергію на відновлення СО2 (хемосинтез).
Денітрифікація - відновлення нітратів до молекулярного азоту чи N2O - відбувається в анаеробних умовах, за яких мікроорганізми використовують для окиснення різних речовин (одержання енергії) кисень нітратів із вивільненням з них азоту. Істотну роль в цих процесах відіграють бактерії р. Pseudomonas. У прісних водах, особливо забруднених, значну роль в цих процесах відіграє кишкова паличка Escherichia coli. Денітрифікація відбувається там, де органічних речовин надходить більше, ніж кисню, необхідного для їхнього біологічного окиснення. Такі умови створюються в гіполімніоні евтрофних озер, в болотах і горизонтах ґрунтових вод зі значним притоком органіки з поверхні.
Амоніфікація - процес розкладу органічних азотистих речовин з виділенням аміаку - відбувається внаслідок руйнування білків організмами. Відбувається в аеробних і анаеробних умовах. Якщо при амоніфікації білків, що містять сірку, утворюються сірководень, індол, скатол, то цей процес називають гниттям.
Амоніфікацію викликають мікроорганізми - амо- ніфікатори: гнильні бактерії, уробактерії, актиноміцети, гриби. Слід підкреслити, що процес амоніфікації притаманний практично всім організмам (реакції дезамінування). Щороку до Світового океану річками виноситься близько 10 млн. т нітрогену в йонній формі та близько 20 млн. т - у формі органічних сполук.8.2.1.2. Колообіг карбогену
Концентрація СО2 в атмосфері складає близько 0,03 % (об’ємних). Загальні запаси його в атмосфері - 711 млрд. т, в осадових породах зосереджено близько 20000000 млрд. т карбогену, 39000 - у водах океану, 12000 - в горючих копалинах. Відносно постійний рівень диоксиду вуглецю в атмосфері підтримується, головним чином, зеленими рослинами та карбо- нат-бікарбонатною системою гідросфери. Атмосфера і океан тісно пов’язані між собою обміном диоксиду вуглецю, що здійснюється через поверхню океану. Щороку близько 100 млрд. т атмосферного двоокису вуглецю розчиняється у воді і приблизно стільки ж його заміщується СО2 з океану. Головний шлях цього колообігу - з СО2 - у живу речовину та зворотній процес - із живої речовини - в СО2:
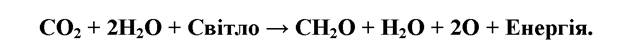
Крім СО2 в атмосфері вуглець представлений також СО - близько 0,1 частина на мільйон, і метаном (СН4) - близько 1,6 частин на мільйон. Як і СО2, ці сполуки знаходяться у стані активного колообігу, час обертання менше місяця для СО і 3,6 року для метану та 4 роки - для СО2. (рис. 8.3).
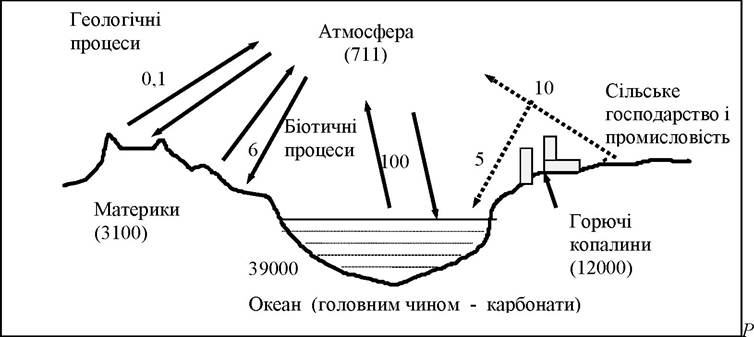
ис. 8.3. Колообіг карбогену в 1015 г (за Ю. Одум, 1986) [18]
Слід відзначити, що метан відіграє важливу роль у підтриманні стабільності озонового шару. Основна кількість метану утворюється у водно- болотних угіддях та на мілководді морів.
Із СО2 значною мірою пов’язаний і парниковий ефект - властивість тропосфери Землі утримувати теплове випромінювання земної поверхні (видиме світло проходить через атмосферу, а інфрачервоні промені, які йдуть від земної поверхні, поглинаються вуглекислим газом (на 64%, метаном - 20%, оксидами азоту - 6% та деякими іншими), що істотно впливає на температурний режим, і саме зі зростанням концентрації СО2 пов’язують підвищення температури на нашій планеті.
Розглянемо баланс СО2 у водних екосистемах. Збагачення води на СО2 відбувається в результаті його інвазії з атмосфери, дихання водних організмів, виділення з різних сполук, в першу чергу з солей вугільної кислоти. Зниження концентрації СО2 відбувається внаслідок евазії в атмосферу, споживання фотосинтетиками та зв’язування в солі вугільної кислоти. Коефіцієнт абсорбції СО2 за температури 00С дорівнює 1,713. Відповідно, за нормального вмісті газу в атмосфері 0,3 мл/л в 1 л води може розчинитися: 0,3 мл ? 1,713 = 0,514 мл СО2.
Зі зростанням температури і солоності нормальний вміст двоокису вуглецю у воді знижується. Частина молекул СО2 реагує з водою, утворюючи вугільну кислоту, яка потім дисоціює:
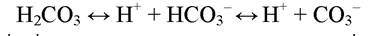
У цій системі співвідношення окремих компонентів залежить від рН середовища. Йони НСО3- і СО32-, реагуючи з іонами металів, утворюють солі, з яких найбільше значення мають карбонати магнію і кальцію. Розчинність CaCO3 незначна і вже при невеликих концентраціях вона випадає в осад. Коли ж вода підкислюється, карбонати перетворюються на розчинні бікарбонати. Розчинення монокарбонатів може тривати до того часу, аж поки весь їхній запас не перейде в бікарбонати. Зворотній процес відбува-
13 7
ється при збільшенні лужності води. Таким чином, у природних водах існує потужна буферна система, що завадить значним коливанням СО2 і рН середовища. Вуглекислота карбонатів називається зв’язаною, а розчинна у воді - вільною.
СО2 є джерелом вуглецевого живлення автотрофів. У незначних кількостях вуглекислота необхідна і тваринам для регуляції метаболізму і синтезу органічних речовин. У результаті реакцій карбоксилювання вуглець включається до складу білків, вуглеводів, ліпідів, нуклеїнових кислот.
За певної (оптимальної) концентрації вуглекислота в середовищі мешкання гідробіонтів є важливою фізіологічно активною речовиною, необхідною для нормального протікання метаболічних процесів, пов’язаних з синтезом основних органічних сполук. Той факт, що підвищення рівня розчиненої у воді вуглекислоти пов’язане з підвищеною її утилізацією в реакціях карбоксилювання, притаманних костистим рибам, безхребетним тощо, має важливе екологічне значення (Романенко та ін., 1980). Дослідження, проведені на водних тваринах, показали існування певного зв’язку між інтенсивністю протікання реакцій карбоксилювання в їх організмі та концентрацією розчиненої у воді вуглекислоти.
8.2.1.2. Колообіг сірки
Сірка присутня у воді у складі сульфатного і сульфідного йонів, у вільному стані, у формі H2S та інших. Із атмосфери сірка в складі тих чи інших сполук потрапляє у водойми. Вихід її з гідросфери пов’язаний, головним чином, з виділенням H2S, зв’язуванням в осадах і біотичним шляхом (вилучення організмів людиною, птахами тощо). У межах водойми коло- обіг сірки зводиться до її відновлення до сірководню в анаеробних умовах і окисненню її за наявності вільного кисню. Обидва процеси пов’язані зі життєдіяльністю організмів.
У прісних водах сірководень утворюється в результаті анаеробного розпаду органічних речовин в ґрунті і придонних шарах води. Інший шлях утворення сірководню - відновлення сульфатів у процесі дисиміляторної сульфатредукції, яка у величезних масштабах здійснюється в морях. Відновлення сульфатів йде за участю сульфатредукуючих бактерій, зокрема прісноводної Desulfovibrio desulphuricans і морської D. aestuarii.
Сульфатредукуючі бактерії, будучи анаеробами і гетеротрофами, використовують сульфати як акцептори водню при метаболічному окисненні (“сульфатне дихання”), подібно тому, як нітрат- і нітритредукуючі бактерії віддають електрони аніонам, що містять кисень, за відсутності вільного О2. Як субстрати, що окислюються, використовуються деякі органічні кислоти і водень. Саме завдяки діяльності сульфатредукуючих бактерій насичена сірководнем водна товща Чорного моря, придонні горизонти Каспійського моря, багатьох інших акваторій. Сірководень утворюється там, де достатньо сульфатів та анаеробні умови.
У морських і континентальних водоймах сірководень окиснюється за наявності кисню частково хімічним шляхом, а переважно - в результаті життєдіяльності різноманітних бактерій. Безбарвні мікроаерофільні сіркобактерії, зокрема нитчасті Beggiatoa, Thiothrix та інші окиснюють сірководень до елементарної сірки, яка осаджується всередині клітин. Подальше окислення її відбувається, коли сірководень у зовнішньому середовищі вичерпується. Енергія, що вивільняється, використовується на відновлення СО2 (хемосинтез). Інший шлях біологічного окислення H2S пов’язаний з життєдіяльністю ряду фотосинтезуючих зелених і пурпурових бактерій, які використовують сірководень як донатор водню.
Дисбаланс окремих складових колообігу сірки, викликаний діяльністю людини (головним чином, спалюванням сірковмісних видів палива, передусім - вугілля у печах ТЕЦ, внаслідок чого утворюється SO2. Окис- нюючись киснем повітря під дією ультрафіолетового випромінювання, він переходить в SO3, котрий, сполучаючись з водяною парою, утворює сульфатну кислоту) є однією з причин «кислотних дощів»
8.2.2. Осадовий цикл. Колообіг фосфору
До елементів осадового циклу належать фосфор, залізо, калій, кальцій, магній та інші елементи, які практично не утворюють газоподібних сполук. Головними рисами осадового циклу є розбалансованість і однонап- равленість. Їх резервний фонд знаходиться в літосфері, а ступінь замкненості цих колообігів у порівнянні з колообігами газоподібних речовин незначний. Розглянемо цей тип колообігу на прикладі фосфору.
Фосфор належить до найбільш дефіцитних елементів біосфери - відношення (вміст у живій речовині)/(вміст у земній корі) для фосфору найвище з біогенних елементів, тому він найчастіше є лімітуючим біопро- дуктивність екосистем фактором.
Колообіг фосфору в біосфері внаслідок нелетючості його сполук не збалансований. Головні запаси його зосереджені в гірських породах, з яких поступово водорозчинні фосфати потрапляють у наземні екосистеми. Внаслідок вимивання фосфати потрапляють у континентальні водойми. Щорічно річками виноситься до Світового океану близько 2 млн. т. фосфору. Зворотній шлях фосфору з океану на суходіл і в континентальні водойми вкрай обмежений і пов’язаний, головним чином, з виловом риби та інших морепродуктів людиною і птахами та з анадромними міграціями риб. Зокрема, враховуючи, що середньобагаторічні величини промислу риби складають близько 70 млн. т/рік, а середній вміст фосфору в тілі риб складає близько 0,8% на сиру вагу (Гандзюра, 1985, 1986), отримуємо 560 000 т, що на порядок (!) більше від величини (60000 т), що цитується в більшості підручників з екології, зокрема Ю. Одума [1.8,] (т. I. с. 211) ! |
У зв’язку з інтенсифікацією сільського господарства все більше фосфору потрапляє до внутрішніх водойм, викликаючи їх евтрофікацію (знач-
не зростання продуктивності, що часто знаходить свій прояв у “цвітінні” водойм тощо).
8.2.3. Колообіг води та його значення
„Вода - візничий природи ” (Леонардо да Вінчі)
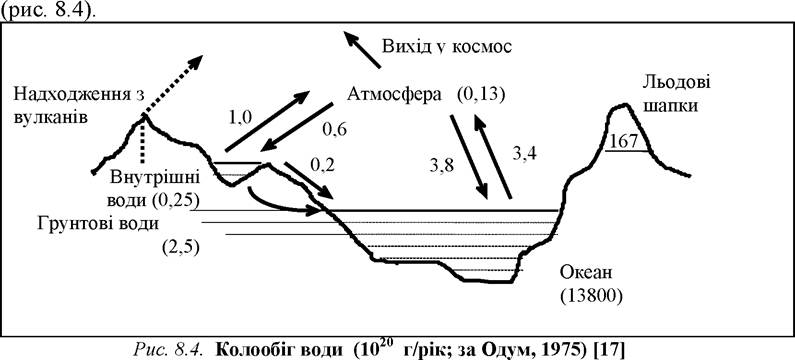
Вода - найпоширеніша речовина у біосфері. Світові запаси води складаються з рідкої (солона і прісна), твердої (прісна) і газоподібної. Загальний об’єм води на нашій планеті сягає 1500 млн. км3. На площу відкритого водного дзеркала (362 млн. км2) припадає близько 71% від загальної площі поверхні Землі (близько 510 млн. км2).
Океани і моря (рідка солона вода) складають близько 97% усієї води. З решти 3% три чверті зберігається у вигляді полярних крижаних шапок і в льодовиках (24 млн. км3). Середній вміст водяного пару в атмосфері відповідає товщині шару води 3 см.
Водна оболонка Землі представлена Світовим океаном, підземними водами і континентальними водоймами, в яких сконцентровано відповідно близько 1370, 6 і 0,23 млн. км3 води.
Під впливом сонячної енергії відбувається безперервний колообіг води. Щорічно з поверхні Світового океану випаровується близько 453 тис. км3 води, із суходолу - 72 тис. км3. Та ж сумарна кількість води (в середньому 525 тис. км3) випадає на Землю у вигляді опадів, але на Океан їх припадає відносно менше, ніж на суходіл (відповідно 411 і 114 тис. км3). Різниця у водному балансі в Світовому океані, що виникає при цьому, поповнюється річковим стоком, що в середньому складає 42 тис. км3 в рік
Територія України з півдня омивається водами Чорного і Азовського морів, які в системі Світового океану займають ізольоване внутрішньоматерикове положення. Берегова лінія Чорного моря в межах України має довжину 1540 км. З берегової лінії Азовського моря, яка простяглася на 2686 км, більше половини знаходиться на території України.
Континентальні водні об’єкти України представлені природними (озера, річки, болота) та штучно створеними водосховищами річкового типу, водоймами-охолоджувачами теплових і атомних електростанцій, ставами рибогосподарського призначення, каналами територіального перекидання водного стоку.
Водні ресурси України є обмеженими і дуже нерівномірно розподіленими за територією. Сумарна величина стоку річок України (без Дунаю) в середній за водністю рік становить 87,1 км3, з них на території країни формується 52,4 км3 води. Щодо водозабезпеченості, то населення України належить до найменш забезпечених водою країн Європи: в Україні воно складає 1,71 (тис. м3 на рік на людину). Схожі величини цього показника у Польщі (1,72) і Румунії (1,76), Чехії і Словаччині (по 1,9). У Норвегії (96,9), Росії (29,6), Швеції (24,1), Фінляндії (22,5), Ірландії (13,7), Латвії (12,2), Естонії (10,1), Білорусії (5,58). Найнижчі значення цей показник має в Угорщині (0,61), Нідерландах (0,78), Люксембурзі (0,94) і Німеччині (1,32).
На території України протікає 63119 річок, які формують рясну мережу водотоків загальною довжиною понад 206 тис. км. Щільність річкової мережі складає 0,25 км на 1 км2 території. За винятком Західного Бугу, який належить до басейну Вісли, всі інші річки належать до басейнів Чорного і Азовського морів.
Усі річки поділяються на три категорії: малі, середні і великі. В основу такого поділу покладено розмір водозбірної площі.
До малих річок належать водотоки, водозбірна площа яких не більша від 2000 км2. При цьому вона повинна розміщуватись в одній геоморфологічній зоні з характерним гідрологічним режимом.
Середні річки мають водозбірну площу 2001--50000 км2. Вони розташовані, як правило, в межах однієї геоморфологічної зони.
На відміну від малих і середніх річок, великі річки охоплюють не одну, а кілька геоморфологічних зон, які можуть істотно відрізнятися за характером водозбірної площі, яка перевищує 50000 км2.
В Україні налічується 63029 малих річок, 81 - середніх і 9 - великих. Великі річки - це Дніпро, Прип’ять, Десна, Дунай, Тиса, Дністер, Південний Буг, Західний Буг, Сіверський Донець. За винятком Десни, Прип’яті, Тиси вони є головними річками відповідних басейнів і мають щільну мережу притоків першого, другого і наступних порядків.
8.3.
Еще по теме Речовинна структура екосистем. Біогеохімічні колообіги:
- Інформаційна структура екосистем
- Енергетична структура екосистем
- Класифікація екосистем
- Жива речовина і її енергія
- Класифікація екосистем
- Нормування забруднюючих речовин у ґрунті
- Нормування забруднюючих речовин у повітрі
- КРУГООБІГ РЕЧОВИН І ХІМІЧНИХ ЕЛЕМЕНТІВ
- Нормування забруднюючих речовин у продуктах харчування
- Роль кліматопу у функціонуванні екосистем
- Нормування забруднюючих речовин у водному середовищі
- Забруднення біосфери та екосистем
- Нормування вмісту забруднюючих речовин у навколишньому середовищі
- Трав’яні типи екосистем