Найважливіші абіотичні фактори та адаптації до них
Величезну кількість абіотичних факторів Г. Вальтер (1979) звів до п’яти первинних прямих (тепло, світло, вода, хімічні, механічні), які мають одиниці виміру і не можуть бути поділені на складові, і чотири групи комплексних (кліматичні, орографічні, едафічні та біотичні), які не мають одиниць виміру.
Нижче розглянемо деякі з них.3.4.1. Тепло
Тепло — це один із найголовніших екологічних чинників, який визначає розподіл організмів на земній кулі. Кількісним показником тепла є температура, яка відображає середню кінетичну швидкість атомів і молекул у будь-якій системі. Від температури залежить і швидкість біохімічних реакцій в організмі, обмін його речовин. Швидкість реакцій визначається коефіцієнтом, який показує, у скільки разів змінюється швидкість реакцій при зміні температури на 10 °C (позначається Q10) і відносно легко розраховується:
Qio = (Al )⅛,
10 XK2)
де K1 і K2 — рівень метаболізму за температури T1 і T2
Для більшості хімічних реакцій величина цього коефіцієнта дорівнює 2—3 (закон Вант-Гоффа). Оскільки величина Q10 для різних біохімічних реакцій різна, зміни температури можуть сильно впливати на збалансованість обміну речовин.
Істотне зниження температури становить небезпеку такого уповільнення обміну речовин, за якого стане неможливим здійснення основних життєвих функцій організму. Критичний момент — замерзання води у клітинах, оскільки поява кристалів льоду несумісна зі збереженням цілісності внутрішньоклітинних структур.
Підвищення температури веде до денатурації білків. Цей процес спостерігається приблизно при +60 °C, але порушення біохімічних і фізіологічних процесів починається раніше, вже при деякому перевищенні +42...+43° C. Посилення метаболізму при високих температурах тіла також може вивести організм із ладу ще задовго до теплового руйнування ферментів, оскільки різко зростають потреби у поживних речовинах і кисні.
Таким чином, життя організмів у середовищі з низькими, високими та мінливими температурами потребує адаптацій, що формуються у процесі еволюції та індивідуального розвитку.У процесі еволюції живих організмів виробилися різноманітні пристосування, які дозволяють регулювати обмін речовин при змінах температури навколишнього середовища. Це досягається двома шляхами: 1) різними біохімічними та фізіологічними перебудовами (зміна набору, концентрації та активності ферментів, зневоднення, зниження точки замерзання розчинів тіла тощо); 2) підтриманням температури тіла на стабільнішому рівні, ніж температура навколишнього середовища, що дозволяє не порушувати хід біохімічних реакцій, що склався.
Температурні межі існування видів
У середньому активна життєдіяльність організмів вимагає досить вузького діапазону температур, обмеженого критичними порогами замерзання води та теплової денатурації білків, приблизно в межах від 0 до +50 °C. Існують екологічні групи організмів, оптимум яких зміщений у бік низьких або високих температур.
Кріофіли — види, що надають перевагу холоду та спеціалізовані до життя в цих умовах. Понад 80 % біосфери сконцентровано в постійно холодних областях із температурою нижче +5 °C — це глибини Світового океану, арктичні та антарктичні пустелі, тундра, високогір’я. Види, що мешкають тут, мають підвищену холодостійкість. Основні механізми цих адаптацій біохімічні — значну роль тут відіграють біохімічні механізми, які запобігають утворенню льоду усередині клітин. При цьому реалізуються два основні шляхи — протистояння замерзанню (резистентність) і стійкість до замерзання (толерантність).
Біохімічний механізм резистентності — накопичення у клітинах макромолекулярних речовин-антифризів, які знижують точку замерзання рідин тіла та перешкоджають утворенню кристалів льоду в організмі. Такого типу адаптації до холоду виявлені, наприклад, у антарктичних риб родини нототенієвих, які живуть за температури тіла —1,86 °C, плаваючи під поверхнею суцільного льоду у воді з такою ж температурою.
Глибоководні риби у приполярних районах також весь час знаходяться у переохолодженому стані.Інший шлях холодостійкості — толерантність — пов’язаний із тимчасовим припиненням активного стану (гіпобіозом або криптобіозом). У комах, наприклад, накопичення захисних органічних речовин, таких як гліцерин, сорбіт, манніт і інших, перешкоджає кристалізації внутрішньоклітинних розчинів і дозволяє переживати критичні морозні періоди у стані заціпеніння. Жуки-туруни у тундрі витримують переохолодження до —35 °C, накопичуючи до зими близько 25 % гліцерину та знижуючи вміст води в тілі з 65 до 54 %. Улітку гліцерину у їх тілі не виявлено. Деякі комахи витримують узимку до —47 і навіть —50 °C із замерзанням позаклітинної, але не внутрішньоклітинної вологи.
Термофіли — це екологічна група видів, оптимум життєдіяльності яких приурочений до області високих температур. Термофілією відрізняються багато представників мікроорганізмів, рослин і тварин, що трапляються в гарячих джерелах, на поверхні ґрунтів, що прогріваються, в органічних залишках, що розкладаються при їх само- розігріванні.
Верхні температурні межі активного життя відрізняються у різних груп організмів. Найбільш термостійкі — бактерії. У деяких видів архебактерій, поширених у воді термальних джерел, експериментально виявлено здатність до росту та поділу клітин за температур, що перевищують +110 °C. Деякі бактерії, що окислюють сульфур (наприклад, Sulfolobus acidocaldarius), розмножуються при +85...+90 °С.
Відомі десятки видів, здатних бути активними при +50 °C і вище в таких місцях існування, як компости, копиці сіна, зерно, що зберігається, ґрунт, що прогрівається, звалища тощо. Найпростіші — амеби та інфузорії, одноклітинні водорості можуть розмножуватися за температури близько +54...+56 °С. Критичні температури тіла деяких тварин, наприклад пустельних ящірок, можуть досягати +48.+49 °С, але для більшості видів температури тіла, що перевищують +43.+44 °С, несумісні з життям унаслідок розбалансування фізіологічних процесів і коагуляції колагену.
З ускладненням організації живих істот їх здатність бути активними при високих температурах знижується.Вузька спеціалізація та латентні стани набагато розширюють межі життя відносно окремих чинників середовища. Якщо середні температурні межі активності організмів характеризуються діапазоном від 0 до +40.+45 °С, то спеціалізовані види (кріофіли та термофіли) розширюють його більше ніж удвічі (від —10 до близько +110 °C), а у стані криптобіозу та анабіозу деякі форми життя здатні витримувати температури, близькі до абсолютного нуля або такі, що набагато перевищують точку кипіння.
Температура тіла і тепловий баланс організмів
Температура тіла живих істот по-різному залежить від температури навколишнього середовища. Баланс тепла в організмі складається з його надходження та витрат. Джерела надходження теплової енергії поділяються на зовнішні та внутрішні. Зовнішнє (екзогенне) тепло організм отримує від нагрітих води, повітря, навколишніх предметів, прямої сонячної радіації. При цьому велику роль відіграють площа покривів та їх теплопровідність. Внутрішнє (ендогенне) тепло виробляється як обов’язковий атрибут обміну речовин.
Для характеристики організмів за основними джерелами необхідного їм тепла використовують терміни «ектотермний» («екзотермний») і «ендотермний». Ектотермія— життєдіяльність переважно за рахунок нагрівання із зовнішнього середовища, ендотермія — за рахунок тепла, що виробляється самим організмом.
Витрати тепла відбуваються через поверхню тіла за рахунок випромінювання та теплопровідності, а також за рахунок енергоємного випаровування води організмами. Співвідношення цих теплообмінних процесів визначає температуру живих істот і впливає на швидкість метаболічних реакцій.
Життєдіяльність і активність більшості видів на Землі залежать перш за все від тепла, що надходить ззовні, а температура тіла зумовлена зовнішньою температурою. Такі організми називають пойкілотермними. Пойкілотермність властива всім мікроорганізмам, грибам, рослинам, безхребетним
тваринам і значній частині хордових.
Дві групи вищих тварин — птахів і ссавців — відносять до гомойотермних. Вони здатні підтримувати постійну оптимальну температуру тіла у широкому діапазоні температур середовища.Серед пойкілотермних організмів є такі, які все життя проводять в умовах постійних зовнішніх температур (глибини океанів, печери тощо), у зв’язку з чим температура їх тіла не змінюється. Таке явище називають удаваною гомойотермією. Вона властива, наприклад, ряду риб і голкошкірих.
Ще одна група тварин — гетеротермні. До неї входять види, що впадають у сплячку або тимчасове заціпеніння. Ці види в активному стані підтримують постійну температуру тіла на високому рівні, а в неактивному — знижену, що супроводжується уповільненням обміну речовин. Це бабаки, ховрахи, кажани, вовчки, їжаки, колібрі, серпокрильці тощо. Таким чином, терміни «пойкілотермія», «гомойотермія», «удавана гомойотермія» і «гетеротермія» відображають ступінь мінливості температури тіла живих істот.
Усі живі організми потенційно ендотермні, але значно відрізняються за рівнем обміну та можливостями збереження тепла. Порушення теплового балансу змінюють температуру тіла. Відновити порушений баланс можна трьома шляхами:
1) зміною теплопродукції, 2) зміною тепловіддачі та 3) переміщенням у просторі у зону оптимальних температур середовища. Пойкілотермні та гомойотермні організми по-різному реалізують можливості температурних адаптацій.
Температурні адаптації пойкілотермних організмів
Температура тіла пойкілотермних тварин змінюється услід за температурою навколишнього середовища. Вони переважно ектотермні: вироблення та збереження власного тепла у них недостатньо для протистояння тепловому режиму місця існування. У зв’язку з цим реалізуються два основні шляхи адаптації: спеціалізація та толерантність.
Спеціалізовані види стенотермні, вони пристосовані до життя у таких ділянках біосфери, де коливання температури відбуваються лише у вузьких межах. Вихід за ці межі для них згубний. Наприклад, коралові поліпи живуть лише в діапазоні температур води від +20,5...+30,0 °C, тобто у тропічному поясі океану.
Голотурія Elpidia glacialis мешкає при температурі води 0...+1 °C і не витримує відхилення від цього режиму ані на градус.Інший шлях адаптації пойкілотермних видів — розвиток пристосувань клітин і тканин до широкого коливання температур. Цей шлях (толерантність) пов’язаний із періодичним гальмуванням обміну речовин і переходом організмів до латентного стану, особливо тоді, коли температура середовища істотно відхиляється від оптимуму.
Ефективні температури розвитку пойкілотермних організмів. Залежність темпів росту та розвитку від зовнішніх температур дає можливість розрахувати проходження життєвого циклу видів у конкретних умовах. Після холодового пригнічення нормальний обмін речовин особин відновлюється для кожного виду за певної температури, яка зветься температурним порогом розвитку, або біологічним нулем розвитку. Чим більше температура середовища перевищує порогову, тим інтенсивніше протікає розвиток і, отже, тим швидше завершується проходження окремих стадій і всього життєвого циклу організму (онтогенезу).
Для здійснення генетичної програми розвитку пойкіло- термним організмам необхідно отримати ззовні певну кількість тепла. Це тепло вимірюється сумою ефективних температур. Під ефективною температурою розуміють різницю між температурою середовища та температурним порогом розвитку організмів. Для кожного виду вона має верхні межі, оскільки дуже високі температури вже не стимулюють, а гальмують розвиток.
Суму ефективних температур розраховують за формулою: X = (T - C) ■ t,
де X — сума ефективних температур, T — температура навколишнього середовища, С — температура порога розвитку і t — кількість годин або діб із температурою, що перевищує поріг розвитку.
Знаючи середній хід температур в будь якому районі, можна розрахувати появу певної фази або кількість можливих генерацій виду, що цікавить нас. У кліматичних умовах Північної України може відродитися лише одна генерація метелика яблуневої плодожерки, а на півдні України — до трьох, що необхідно враховувати під час розроблення заходів захисту садів від шкідників. Саме сума ефективних температур, яку потрібно набрати для завершення життєвого циклу, часто обмежує географічне поширення видів.
Температурна компенсація. Ряд пойкілотермних видів, що мешкають в умовах мінливих температур, мають здатність підтримувати більш-менш постійний рівень обміну речовин у досить широких межах зміни температури тіла. Це явище зветься температурною компенсацією і відбувається в основному за рахунок біохімічних адаптацій. Наприклад, у черевоногих (літторини, Littorina Iittorea) і двостулкових (мідій, Mytilus edulis) молюсків, які трапляються на узбережжі Баренцового моря, інтенсивність обміну речовин майже не залежить від температури в тих межах, з якими молюски стикаються щодня під час припливів і відпливів. У весняно-літній період цей діапазон сягає понад 20 °C (від +6 до +30 °C). У холодній воді їх метаболізм такий же інтенсивний, як і в теплому повітрі. Це забезпечується роботою ферментів, які при пониженні температури змінюють свою конфігурацію таким чином, що зростає їх спорідненість до субстрату, і реакції протікають активніше.
Інші способи температурної компенсації пов’язані із заміною ферментів, що діють, на подібні за функцією, але які працюють при іншій температурі (ізоферменти). Такі адаптації вимагають часу, оскільки відбувається інактивація одних генів і вмикання інших із подальшими процесами побудови білків. Подібна аклімація (зміщення температурного оптимуму) лежить в основі сезонних перебудов, а також виявляється у представників поширених видів у різних за кліматом частинах ареалу. Ряд пойкілотермних організмів має здатність до часткової регуляції теплообміну, тобто певними способами збільшувати надходження тепла до організму або віддавати його надлишок.
Можливості регуляції температури у пойкілотермних тварин. Найважливіша особливість тварин — їхня рухливість, здатність переміщатися у просторі — створює принципово нові адаптивні можливості, зокрема й у терморегуляції. Тварини активно обирають місце життя зі сприятливішими для них умовами. На відміну від рослин тварини, що мають мускулатуру, продукують значно більше власного, внутрішнього тепла. Під час скорочення м’язів вивільняється значно більше теплової енергії, ніж при функціонуванні будь-яких інших органів і тканин, оскільки ККД використання хімічної енергії для здійснення м’язової роботи відносно низький. Чим потужніша та активніша мускулатура, тим більше тепла може генерувати тварина.
Пойкілотермні тварини залишаються екзотермними, оскільки загальний рівень їх метаболізму не настільки високий, щоб внутрішнього тепла було достатньо для обігріву тіла. Проте деякі з пойкілотермних тварин у стані активності здатні підтримувати температуру тіла вищу, ніж у навколишньому середовищі. Наприклад, метелики-бражники, що ведуть нічний спосіб життя, літають і живляться на квітках навіть при +10 °C. Під час польоту температура їх грудного відділу підтримується на рівні +40...+41 °C. Інші комахи можуть літати у холодному повітрі, заздалегідь розігріваючи свої літальні м’язи для злету (сарана, джмелі, оси, бджоли, великі нічні совки). Джмелі збирають нектар навіть при +5 °C, маючи температуру тіла +36...+38 °C. При припиненні активності комахи швидко остигають. Генерувати тепло для обігріву можуть у деяких випадках і рептилії. Самка пітона, що обвиває своїм тілом кладку, скорочуючи мускулатуру, здатна
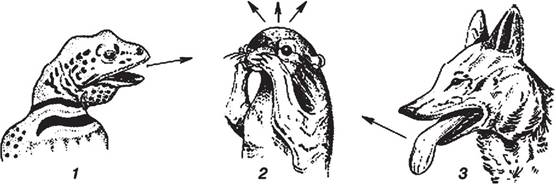
Рис. 3.8. Випаровувальна терморегуляція у тварин (Чернова, Би- лова, 2004): 1 — випаровування зі слизових оболонок (ящірка), 2 — натирання слиною (антилоповий ховрах), 3 — випаровування за рахунок частого дихання (собака)
підвищувати температуру на 5—6 °C у діапазоні зовнішніх температур +25...+33 °C. У прохолоднішому повітрі змія стає млявою та неактивною.
Основні способи регуляції температури тіла у пойкіло- термних тварин — поведінкові: зміна пози, активний пошук сприятливих місць, низка спеціалізованих форм поведінки, спрямована на створення мікроклімату (риття нір, спорудження гнізд тощо). Зміною пози тварина може посилити або послабити нагрівання за рахунок сонячної радіації. Наприклад, пустельна сарана у прохолодні ранішні години підставляє сонячним променям широку бічну поверхню тіла, а опівдні — вузьку спинну. Ящірки, навіть високо в горах, у період нормальної активності можуть підтримувати температуру тіла завдяки нагріванню прямими сонячними променями, а також використовуючи тепло нагрітих скель і каміння. У сильну спеку тварини ховаються у тінь, у нори, щілини. У пустелях вдень, наприклад, деякі види ящірок і змій піднімаються на кущі або зариваються в менш нагріті глибші шари піску, уникаючи контактів із розпеченою поверхнею ґрунту. Ще складніші форми поведінки соціальних комах: бджіл, мурашок, термітів, які будують гнізда з добре регульованою усередині них температурою, майже постійною у період їх активності.
У деяких пойкілотермних тварин ефективно діє механізм випаровувальної терморегуляції (рис. 3.8). За рахунок цього жаби за годину при +2θ °C витрачають на суходолі близько 7770 Дж, що у 300 разів більше за їх власну теплопродукцію. Багато рептилій при наближенні температури до верхньої критичної починають важко дихати або тримати рот відкритим, посилюючи віддачу води зі слизових оболонок. Бджоли, що літають у спеку, уникають перегріву, виділяючи з рота краплю рідини, випаровування якої видаляє надлишок тепла.
Незважаючи на ряд можливостей фізичної та поведінкової терморегуляції, пойкілотермні тварини можуть здійснювати її лише у вузькому діапазоні температур. Унаслідок низького загального рівня метаболізму вони не можуть забезпечити постійність теплового балансу і достатньо активні тільки у вузькому діапазоні верхніх температурних меж існування. Мешкання в умовах постійно низьких температур для холоднокровних тварин утруднене.
Температурні адаптації гомойотермних організмів
Гомойотермія — принципово інший шлях температурних адаптацій, що виник на основі різкого підвищення рівня окис- нювальних процесів у птахів і ссавців у результаті еволюційного вдосконалення кровоносної, дихальної та інших систем органів.
Основні відмінності гомойотермних тварин від пойкіло- термних:
1) потужний потік внутрішнього, ендогенного тепла;
2) розвиток цілісної системи ефективних терморегуляторних механізмів;
3) постійне протікання всіх фізіологічних процесів в оптимальному температурному режимі.
Гомойотермні тварини зберігають постійний тепловий баланс між теплопродукцією та тепловіддачею і, відповідно, підтримують постійну високу температуру тіла. Організм типової гомойотермної (не гетеротермної) тварини не може бути тимчасово «зупинений» так, як це відбувається при гіпобіозі або криптобіозі у пойкілотермних.
Гомойотермні тварини завжди забезпечують певний мінімум теплопродукції, що дає можливість роботи кровоносної системи, органів дихання, виділення та інших, навіть у стані спокою. Цей мінімум отримав назву базального метаболізму. Перехід до активності посилює вироблення тепла і, відповідно, вимагає посилення тепловіддачі.
Теплокровним властива хімічна терморегуляція — рефлекторне збільшення теплопродукції у відповідь на зниження температури середовища. Хімічна терморегуляція повністю відсутня у пойкілотермних, у яких, у разі виділення додаткового тепла, воно генерується за рахунок безпосередньої рухової активності тварин.
При дії холоду в організмі теплокровних тварин окис- нювальні процеси не слабшають, а посилюються, особливо у скелетних м’язах. У багатьох тварин спочатку спостерігається м’язове тремтіння — неузгоджене скорочення м’язів, що приводить до виділення теплової енергії. Крім того, клітини м’язової та багатьох інших тканин виділяють тепло і без здійснення робочих функцій, переходячи до стану особливого то- нусу терморегуляції. При подальшому зниженні температури середовища тепловий ефект тонусу терморегуляції зростає.
Під час продукування додаткового тепла суттєво посилюється обмін ліпідів, оскільки нейтральні жири містять основний запас хімічної енергії. Внаслідок цього жирові запаси тварин забезпечують кращу терморегуляцію. Ссавці мають навіть спеціалізовану буру жирову тканину, під час окиснен- ня якої вся хімічна енергія, що вивільняється, розсіюється у вигляді тепла, тобто йде на обігрів організму. Бура жирова тканина найбільш розвинена у тварин — мешканців холодного клімату та гетеротермних тварин.
Підтримання температури тіла за рахунок зростання теплопродукції вимагає значних витрат енергії, тому тварини при посиленні хімічної терморегуляції або мають потребу у великій кількості корму, або витрачають багато жирових запасів, накопичених раніше. Наприклад, бурозубка-крихітка (Sorex minutissimus) має виключно високий рівень метаболізму. У тварини чергуються дуже короткі періоди сну та активності, вона діяльна у будь-яку годину доби і за добу з’їдає корму учетверо більше маси власного тіла. Так само і птахам, що залишаються на зиму у помірних широтах, потрібно багато корму: їм страшні не стільки морози, скільки нестача кормів. Зокрема, при гарному врожаї насіння ялини та сосни шишкарі взимку навіть виводять пташенят. Посилення хімічної терморегуляції, таким чином, має свої межі, обумовлені можливістю добування їжі. При нестачі корму взимку такий шлях терморегуляції екологічно невигідний.
У межах відповідного діапазону зовнішніх температур гомойотермні тварини підтримують температуру тіла, не витрачаючи на це додаткової енергії, а використовуючи ефективні механізми фізичної терморегуляції, що дозволяє краще зберігати або відводити тепло базального метаболізму. Цей діапазон температур, у межах якого тварини відчувають себе найкомфортніше, називається термонейтральною зоною. За нижнім порогом цієї зони починається хімічна терморегуляція, за верхнім — витрати енергії на випаровування.
Фізична терморегуляція екологічно вигідна, оскільки адаптація до холоду здійснюється не за рахунок додаткового вироблення тепла, а за рахунок збереження його в тілі тварини. Крім того, цей вид терморегуляції дає можливість захисту від перегріву шляхом посилення тепловіддачі до зовнішнього середовища.
Способи фізичної терморегуляції численні. У філогенетичному розвитку ссавців механізми фізичної терморегуляції стають все досконалішими та різноманітнішими. До них слід віднести рефлекторне звуження та розширення кровоносних судин шкіри, що міняє її теплопровідність, зміну теплоізолю- вальних властивостей хутра та пір'яного покриву, протиточ- ний теплообмін шляхом контакту судин при кровопостачанні окремих органів, регуляцію випаровувальної тепловіддачі.
Густе хутро ссавців, пір’яний і, особливо, пуховий покрив птахів дозволяють зберігати навколо тіла прошарок повітря з температурою, близькою до температури тіла тварини, і, тим самим, зменшувати випромінювання тепла у зовнішнє середовище. Тепле зимове хутро ссавців Заполяр’я дозволяє їм у холоди обходитися без істотного підвищення обміну речовин і знижує потребу в живленні — песці на узбережжі Північного Льодовитого океану узимку споживають корму навіть менше, ніж влітку. У морських ссавців (ластоногих і китів) шар підшкірної жирової клітковини розподілений по всьому тілу. Товщина підшкірного жиру в особин окремих видів тюленів сягає 7—9 см, а загальна його маса становить до 40—50 % маси тіла. Теплоізолювальний ефект такої «жирової панчохи» настільки високий, що під тюленями, які годинами лежать на снігу, сніг не тане, хоча внутрішня температура тіла тварини підтримується на рівні +38 °C.
Важливе значення для підтримання температурного балансу має відношення поверхні тіла до його об’єму, оскільки відомо, що масштаби продукування тепла залежать від маси тварини, а теплообмін відбувається через її покриви.
Зв’язок розмірів і пропорцій тіла тварин із кліматичними умовами їх мешкання був виявлений ще в XIX ст. Згідно з правилом Бергмана (1848), якщо два близькі види теплокровних тварин відрізняються розмірами, то більший мешкає в холоднішому, а дрібніший — у теплому кліматі. Бергман підкреслював, що ця закономірність виявляється лише в тому випадку, якщо види не відрізняються іншими пристосуваннями до термо - регуляції.
Д. Аллен у 1877 р. помітив, що у багатьох ссавців і птахів Північної півкулі відносні розміри кінцівок та інших виступаючих частин тіла (хвостів, вух, дзьобів) збільшуються з поширенням на південь. Ця закономірність отримала назву правила Аллена. Значення окремих ділянок тіла в терморегуляції далеко не рівноцінне. Виступаючі частини тіла мають велику відносну поверхню, яка вигідна в умовах спекотного клімату. У певних ссавців, наприклад, значення для підтримки теплового балансу мають вуха, насичені великою кількістю кровоносних судин. Величезні вуха африканського слона, маленької пустельної лисички-фенека, американського зайця перетворилися на спеціалізовані органи терморегуляції (рис. 3.9).

Рис. 3.9. Вуха африканського довговухого тушканчика як орган терморегуляції (Соколов и др., 1977)
Під час адаптацій тварин до холоду проявляється закон економії поверхні: компактна форма тіла з мінімальним відношенням площі до об'єму найвигідніша для збереження тепла.
Поведінкові способи регуляції теплообміну для теплокровних тварин не менш важливі, ніж для пойкілотермних: від зміни пози та пошуків схованок до спорудження складних нір, гнізд, здійснення ближніх і дальніх міграцій. У норах рийних тварин хід температур згладжений тим сильніше, чим більша глибина нори. У середніх широтах на глибині 150 см від поверхні ґрунту сезонні коливання температури майже не спостерігаються.
У ряді випадків гомойотермні тварини використовують із метою терморегуляції групову поведінку. Наприклад, деякі пінгвіни в лютий мороз і бурани збиваються у щільну купу, так звану «черепаху». Особини, що опинилися на периферії, через деякий час пробиваються всередину, і «черепаха» поволі кружляє та переміщується. Усередині такого скупчення температура підтримується близько +37 °C навіть у найлютіші морози. Мешканці пустель — верблюди — у спеку також збиваються разом, притискаючись один до одного боками, але цим досягається протилежний ефект — запобігання сильному нагріванню усієї поверхні тіла сонячними променями. Температура у центрі скупчення тварин дорівнює температурі їх тіла (+39 °C), тоді як шкіра на спині та боках крайніх тварин нагрівається до +70 °C.
Поєднання ефективних способів хімічної, фізичної та поведінкової терморегуляції при загальному високому рівні окислювальних процесів в організмі дозволяє гомойотермним тваринам підтримувати свій тепловий баланс на тлі широких коливань зовнішньої температури.
Екологічні вигоди пойкілотермії та гомойотермії
Пойкілотермія та гомойотермія — дві альтернативні стратегії виживання організмів в умовах температур, що змінюються. Кожна з них має свої переваги та недоліки.
Пойкілотермність — підпорядкування організмів перебігу зовнішніх температур. Виробляючи порівняно мало ендогенного тепла та маючи лише окремі терморегуляторні реакції, пойкілотермні організми не можуть забезпечити сталості теплообміну, тому при коливаннях температури середовища їх активність переривчаста. Перехід особин до неактивного стану пов’язаний із розвитком механізмів толерантності їх до змін температури тіла.
Гомойотермність — стратегія протистояння впливу чинників середовища. Організм гомойотермної тварини завжди функціонує тільки у вузьких температурних межах. За цими межами життя неможливе, оскільки організм втрачає здатність витримувати значні коливання температури тіла. Проте, відрізняючись високою інтенсивністю окислювальних процесів в організмі та маючи потужний комплекс терморегуляторних засобів, гомойотермні тварини можуть підтримувати для себе постійний температурний оптимум навіть при значних відхиленнях зовнішніх температур, що дозволяє їм освоювати різні середовища існування.
Поєднання елементів різних стратегій
Ряд тварин здатні поєднувати переваги обох стратегій теплообміну. У деяких видів гомойотермних, що освоюють екстремальні умови, а особини перебувають на межі можливого підтримання теплового балансу тіла, виникають елементи пристосування, що нагадують пойкілотермію і дозволяють економити енергію. Гетеротермія — особлива адаптивна стратегія птахів і ссавців, коли закономірно поєднується використання вигод як постійності температури тіла, так і її зміни. Основні форми прояву гетеротермії — здатність впадати в сплячку або торпідний стан (заціпеніння).
Зимівля характерна для низки ссавців, поширена у однопрохідних, комахоїдних, гризунів і рукокрилих. Впадаючи у сплячку, тварини припиняють підтримання високої температури тіла, знижуючи її лише до декількох градусів вище нуля. Часто тварини проводять у сплячці довгі місяці (іноді до половини року), поступово витрачаючи накопичені резерви (табл. 3.1). Зимовий сон ведмедів, до речі, не називають сплячкою, оскільки температура цих звірів знижується лише на 3—6 °C, а зниження рівня метаболізму незначне.
Торпідний стан, або заціпеніння, супроводжує зимівлю тварин, але виникає і в інших умовах як самостійна адаптація. Низка дрібних видів птахів і ссавців знижує рівень обміну і температуру тіла за несприятливих погодних змін або регулярно в добових циклах. У колібрі, наприклад, із настанням ночі температура тіла знижується з +36...+40 до + 18 °C, птахи сідають на гілки й ціпеніють. Кажани, навпаки, активні у присмеркові та нічні години і впадають у торпід- ний стан удень. Перехід до заціпеніння стимулюється також і нестачею корму. За одних і тих самих температурних умов забезпечені кормом тварини можуть продовжувати активний спосіб життя й генерувати тепло на терморегуляцію, а за браку їжі впадають у стан заціпеніння.
Таблиця 3.1.
Кратність зниження рівня метаболізму під час сплячки (Мс) порівняно з активним станом (Ма) у гризунів (Шилов, 1998)
| Види | Маса тіла, г | Мс/Ма |
| Байбак альпійський (Marmota marmota) | 2007 | 21,5 |
| Їжак звичайний (Erinaceus europaeus) | 642 | 46,6 |
| Ховрах європейський (Citellus citellus) | 227 | 53,0 |
| Вовчок (Glis glis) | 129 | 72,6 |
| Соня садова (Eliomys quercinus) | 63 | 56,6 |
| Соня ліщинова (Muscardinus avellanarius) | 19 | 76,6 |
Між короткочасним заціпенінням гетеротермних тварин і тривалою зимівлею існує низка переходів. У пустелях ряд дрібних ссавців (мишоподібні гризуни, вовчки, деякі ховрахи, тенреки тощо) впадають не тільки у зимову, а і в літню сплячку, яка зветься естивацією (від лат. aestes — літо). Температура їх тіла падає при цьому не так значно (до +25...+27 °C), але відповідне зниження рівня метаболізму дозволяє економити не тільки енергію, а й воду. Таким чином, широке поширення гетеротермності дозволяє вважати фундаментальним значення цього явища для виживання тварин у несприятливих умовах.
3.4.2. Світло
Усім живим організмам для життєдіяльності необхідна енергія, що надходить ззовні. Основне джерело її — сонячна радіація, на яку припадає близько 99,9 % загального балансу енергії Землі. Якщо прийняти сонячну енергію, що досягає Землі, за 100 %, то приблизно 19 % її поглинається під час проходження через атмосферу, 34 % відбивається у космічний простір і 47 % досягає земної поверхні у вигляді прямої та розсіяної радіації. Пряма сонячна радіація — континуум електромагнітного випромінювання з довжинами хвиль від 0,1 до 30 000 нм. На ультрафіолетову частину спектра припадає 1—5 %, на видиму — 16—45 %, на інфрачервону — 49—84 %.
Розподіл сонячної енергії за спектром істотно залежить від маси атмосфери й змінюється залежно від висоти стояння Сонця. Кількість розсіяної радіації (відбиті промені) зростає зі зменшенням висоти стояння Сонця та збільшенням каламутності атмосфери. Спектральний склад сонячної радіації безхмарного неба характеризується максимумом енергії в діапазоні 400—480 нм.
Світло як умова орієнтації тварин
Для тварин сонячне світло не є таким вкрай необхідним чинником, як для зелених рослин, оскільки всі гетеротрофи існують за рахунок енергії, накопиченої автотрофами. Проте й у житті тварин світлова частина спектра сонячного випромінювання відіграє важливу роль. Різні види тварин мають потребу у світлі певного спектрального складу, інтенсивності та тривалості освітлення. Відхилення від норми пригнічують їх життєдіяльність і навіть призводять до загибелі. Розрізняють види світлолюбні (фотофіли) та тіньолюбиві (фотофоби); еврифотні таксони витримують широкий діапазон освітленості, стенофотні — обмежені умови освітленості.
Світло для тварин — необхідна умова бачення, зорової орієнтації у просторі. Розсіяні, відбиті від навколишніх предметів промені, сприйняті органами зору тварин, дають їм значну частину інформації щодо зовнішнього світу. Повнота зорового сприйняття навколишнього середовища залежить у тварин у першу чергу від ступеня еволюційного розвитку. Примітивні вічка багатьох безхребетних — просто світлочутливі клітини, оточені пігментом, а в одноклітинних — світлочутлива ділянка цитоплазми. Органи зору з окремих вічок не дають зображення предметів, а сприймають лише коливання освітленості, чергування світла та тіні, що свідчать про зміни навколишнього середовища. Павуки, наприклад, можуть розрізняти контури рухомих предметів на відстані 1—2 см.
Образне бачення можливе лише за достатньо складної будови ока. Найдосконаліші органи зору — очі хребетних, головоногих молюсків і комах. Вони дозволяють сприймати форму та розміри предметів, їх колір, визначати відстань до них.
Поняття видимого світла певною мірою умовне, оскільки окремі види тварин сильно різняться за здатністю сприймати різні частини сонячного спектра. Деякі тварини, наприклад гримучі змії, відчувають інфрачервону частину спектра та ловлять здобич у темряві, орієнтуючись за допомогою спеціальних органів. Для бджіл видима частина спектра зміщена до короткохвильової зони. Вони сприймають як колір значну частину ультрафіолетових променів, але не розрізняють червону частину спектра.
Здатність до розрізнення кольорів значною мірою залежить і від того, за якого спектрального складу випромінювання активні особини виду. Більшість ссавців, що походять від предків із присмерковою та нічною активністю, погано розрізняють кольори і бачать усе у чорно-білому зображенні (собачі, котячі, хом’яки тощо). Такий же зір характерний для нічних птахів (сови, дрімлюги). Денні птахи мають добре розвинений кольоровий зір.
Життя за присмеркового освітлення часто призводить до гіпертрофії очей. Величезні очі, здатні вловлювати незначні відсотки світла, властиві лемурам, лорі, довгоп’ятам, совам та іншим тваринам, що ведуть нічний спосіб життя.
Проживання в умовах повної темряви, як правило, пов’язане з редукцією органів зору. Зокрема це властиве видам, особини яких мешкають у печерах, а також багатьом ґрунтовим тваринам. Утім, в останніх нерідко є світлочутливі органи, хоча й у редукованому вигляді, все ж зберігаються та використовуються для отримання інформації про вихід на освітлену поверхню.
В океані інтенсивність освітлення знижується з глибиною, змінюється також і його спектральний склад: найглибше проникає короткохвильова частина сонячних променів — синього та блакитного кольорів. Освітленість на мілководді мало відрізняється від суходолу. Риби, що мешкають тут, мають у сітківці значний відсоток рецепторів (колбочок), чутливих до червоного кольору. У риб, що мешкають у зеленій воді прибережної зони, таких колбочок немає; відсутні у них і чутливі до помаранчевого кольору клітини.
Серед глибоководних риб більшість має в сітківці лише один тип паличок, чутливих до синього кольору. На глибині 800—950 м інтенсивність світла складає близько 1 % освітлення на поверхні. Цього ще достатньо для світосприйняття: поріг зорової чутливості деяких організмів наближається до 10—10 полудневого освітлення. Подальше збільшення глибини пов’язане в одних видів із редукцією органів зору, а в інших — із розвитком гіпертрофованих очей, здатних сприймати навіть дуже слабке світло. Це значною мірою визначається наявністю на великих глибинах організмів, що світяться. Деякі з них здатні створювати освітлення порядку 10—2 мкВт/см2, що близько до порога світлової чутливості тварин. Таке світіння з довжиною хвилі 400—500 нм характерне для блакитного діапазону і відповідає «налаштуванню» органів зору глибоководних тварин. Біологічне світіння використовують і риби, утворюючи симбіотичні зв’язки з мікроорганізмами, що світяться, і формуючи спеціальні органи, світло яких використовується для того, щоб підманювати здобич, для взаємного розпізнавання, розпізнавання статі тощо (рис. 3.10).
Світло та біологічні ритми
Специфічне значення світлового чинника полягає в тому, що закономірна динаміка умов освітлення відіграє важливу роль у регуляції періодичних явищ у житті представників органічного світу. Із самого виникнення життя на нашій планеті

Рис. 3.10. Глибоководні вудильники (Нікольський, 1974): 1 — самка церації (Ceratias holboelli) із прикріпленим самцем (1а); 2 — неоце- рація (Neoceratias spinifer) із прикріпленим самцем (2а); 3 — кау- лофрина (Caylophrine jordani); 4 — меланоцет (Melanocetus apogon); 5 — борофрина (Borophrine apagon); 6 — гігантактис (Gigantactis macronema)
воно розвивалося в умовах ритмічно мінливого середовища. Закономірна зміна дня та ночі, сезонні зміни комплексу чинників, що регулярно повторюються, — все це вимагало пристосування живих організмів. Кардинальна форма такого пристосування виражається в еволюційному становленні співмірності та узгодження ритмів біологічної активності різних живих форм із масштабами добової та сезонної цикліки комплексу умов середовища. Адаптивний сенс цього явища полягає в тому, що на його основі з’явилася можливість поєднати різні форми життєдіяльності організму з найсприятливішим періодом зовнішніх умов. Аналогічним чином в еволюції низки видів формувалися ритми з іншою періодичністю — наприклад місячною або припливно-відпливною.
Ритмічність загальних проявів життєдіяльності та окремих її форм властива всім живим істотам. У її основі лежить специфіка біохімічних і фізіологічних реакцій, що мають ритмічний характер. Тривалість ритмів окремих процесів, що відбуваються на суборганізменому рівні, дуже різна: від часток секунди (наприклад, активність нейрона) до декількох годин (секреторна діяльність залоз) і навіть більше. Функціонування цілого організму базується на інтеграції окремих суборганізмених ритмів і узгодженні їх з умовами середовища, що змінюються в часі.
«Подвійний» характер походження адаптивних циклів (хіміко-біологічна природа первинних ритмів і залежність їх від періодичних змін умов середовища) виразно проявляється у фізіологічних механізмах, які регулюють добову та сезонну періодичність життєдіяльності організмів. За сучасними уявленнями в основі періодичних процесів лежить внутрішня (ендогенна) програма, на яку впливає складний комплекс зовнішніх умов. Одні з цих умов прямо модифікують ендогенну програму відповідно до конкретної екологічної ситуації, інші виступають як «датчики часу», сприяючи синхронізації ендогенних циклів із закономірною (добовою, сезонною) зміною зовнішніх умов. Як «датчики» часу можуть виступати багато періодично змінних факторів середовища. Але в еволюції більшості груп живих організмів основне синхронізувальне значення закріпилося за закономірними змінами світлового режиму — фотоперіодична регуляція.
Світло — первинно-періодичний чинник. Закономірна зміна дня та ночі, як і сезонні зміни тривалості світлої частини доби, відбуваються з чіткою ритмічністю, яка визначається астрономічними процесами і на прояви якої не можуть вплинути умови та процеси, що відбуваються на Землі. Тому фотоперіод найстійкіший у своїй динаміці, автономний і не схильний до інших впливів. Фотоперіодом називають співвідношення світлої та темної частин доби. У більш спеціальному сенсі цей термін використовують для позначення довжини дня («короткий» або «довгий» фотоперіод). Тільки на екваторі, де тривалість дня та ночі не змінюється залежно від сезону, й у деяких особливих умовах (глибини морів, печери, безперервний полярний день) провідного значення в регуляції біологічних ритмів набувають інші чинники.
Добові ритми. Добова періодичність властива більшості тварин. Є форми із денною або нічною активністю, у деяких видів піки активності виявляються спонтанно, незалежно від часу доби, деяким тваринам властивий прояв активності у присмерковий час. Час початку або закінчення неспання (або, навпаки, сну) у тварин є видоспецифічним і відрізняється суттєвою сталістю за відношенням до добової зміни освітленості.
З еволюційної точки зору формування в особин видового стереотипу активності — складний процес, що відображає їх пристосування до багатьох чинників середовища. При цьому для різних тварин умови освітлення далеко не завжди мали пряме відношення до цього процесу. Лише у форм із провідним значенням зорової рецепції (наприклад, птахи) денний тип активності прямо пов’язаний зі світлом як чинником бачення. Але навіть у цьому випадку такі умови, як трофічна конкуренція або спеціалізація живлення, призводили до виникнення окремих форм нічної активності.
Загальний характер активності тварин у більшості випадків визначається такими умовами, як тип живлення, взаємини з хижаками та конкурентами, добові зміни комплексу абіотичних чинників. Так, добова активність пойкілотермних тварин багато в чому визначається режимом температури середовища, хоча для амфібій характерний чинник добової активності — поєднання температури та вологості. Серед гризунів види, що поїдають грубі, багаті клітковиною корми, відрізняються переважно цілодобовою активністю. На- сіннєїдні ж форми, які споживають більш концентрований корм, мають можливість приурочити час його добування до нічного періоду, коли слабшає прес хижаків. Експериментально встановлено, що присмерково-нічний тип активності куницевої акули (Mustelus canis) пов’язаний з аналогічним типом активності її основної їжі — ракоподібних.
Циклічні зміни загального рівня життєдіяльності протягом доби пов’язані з відповідними ритмами фізіологічних процесів. Активний період характеризується суттєвими енер- говитратами та відповідно підвищеною активністю комплексу фізіологічних реакцій. Разом із цим, добові коливання метаболізму не є лише прямим наслідком підвищення загальної активності: існують і закономірні добові зміни рівня обміну спокоєм. Режим освітлення виступає в ролі сигнального чинника, який визначає час початку та закінчення активності.
Порогові величини освітленості визначають час початку та закінчення активності. Разом із цим, упродовж активної частини доби інтенсивність діяльності тварин зазвичай має пульсуючий, фазовий характер. Так, горобині птахи у період розмноження найактивніші в ранішній час, удень їх активність дещо знижується і знову підвищується увечері. Те ж саме характерне і для багатьох нічних видів: сіра сова, наприклад, найактивніша на початку та наприкінці ночі (іноді виявляється третій пік, у середині ночі). Нерівномірний прояв активності пов’язаний із темпами накопичення та витрачання енергетичних ресурсів, пристосуванням до впливу несприятливих чинників (наприклад, перерва активності у спекотний час доби) і тому подібне.
Циркадіанні (циркадні) ритми. Сигнальна, синхронізуюча роль фотоперіоду виразно виявляється в умовах експерименту, коли на фоні незмінної освітленості (найчастіше — у темряві) у піддослідних організмів виявляється добовий ритм, властивий даному виду у природних умовах. Це явище було уперше виявлене й описане ще у XVIII ст. у дослідах із рослинами, які в нормі опускають або складають листя на ніч і розпрямляють удень. Після переміщення у повну темряву ці рослини зберігали добовий ритм руху листя. Таким чином, уже на початок позаминулого століття були показані два фундаментальних принципи сучасної концепції механізмів добової ритміки: наявність ендогенної програми з періодом близько доби та можливість впливу умов освітлення (фотоперіоду) на реалізацію цієї програми.
Ендогенний ритм має дві фази тривалістю близько 12 годин кожна: світлова (світлолюбна) та темнова (темнолюбна). Упродовж цих фаз реакція організму на вплив світла полярно міняється. До такої періодики пристосований і рівень основного обміну, що свідчить про ендогенний характер циклів метаболізму та температури тіла. Широко відома чітка добова періодика мітотичної активності клітин, яка, як правило, знаходиться у протифазі із циклом загальної активності, що, можливо, відображає «раціональний» розподіл енерговитрат організму протягом доби. Так, експерименти з мадрепоровим коралом Acropora acuminata показали, що мічений Карбон включається в їх тканині тільки у денний час із максимумом у післяполудневі години. Інтенсивність видимої кальцифіка- ції максимальна опівдні та мінімальна опівночі. Досліди зі слимаками Laevicaulis alta виявили чітку періодичність рівня фосфорилазної активності з максимумом о 0-й годині та мінімумом о 12-й годині.
У птахів і ссавців відомі добові цикли ряду ендокринних залоз і ферментних систем. Наприклад, у лабораторних мишей за активністю ферментів орнітинового циклу встановлений чіткий добовий ритм метаболізму нітрогену з максимумом о 21-й годині та мінімумом о 9-й годині. Загалом у ссавців відомо не менше 50 органів, що мають власний ендогенний ритм функціонування.
Механізми циркадіанних ритмів і їх регуляція залишаються предметом інтенсивних досліджень. Природа «біологічного годинника», що лежить в основі ендогенних ритмів, вивчена ще недостатньо. Суттєва автономність ритмів в окремих органах, тканинах і клітинах спонукає шукати ці механізми на субклітинному рівні. Ще наприкінці 1940-х років професор Московського університету Д. А. Сабінін висловив припущення про зв’язок механізмів добової фізіологічної періодики зі структурою генетичного апарату. Наприкінці 1960-х років ця гіпотеза була відроджена у вигляді концепції хронона. Згідно з цією концепцією, матеріальним носієм відліку часу служить довга молекула ДНК, нитки якої розходяться, і на них будується інформаційна (матрична) РНК, яка досягає повної довжини одинарної нитки ДНК приблизно за 24 години.
Останнім часом (починаючи з 1970-х років) вважають, що молекулярний механізм циркадіанної ритмічності описується трансляційно-мембранною моделлю, яка поєднує два етапи: трансляції специфічних білків на рибосомах і подальшого вбудовування цих білків у клітинні мембрани. Існує гіпотеза, що у функціонуванні біологічного годинника задіяні дві складові: гіпоталамус і хронон. Гіпоталамічна регуляція реалізується нейросекреторною системою і пов’язується з участю гуморальних механізмів. Показано, зокрема, що у ссавців циркадіанний ритм мітозів і вмісту еозинофілів у периферичній крові регулюється з участю надниркових залоз, а клітинні цикли — на основі моделі хронона.
У середині минулого століття ці принципи отримали підтвердження в численних експериментах із тваринами різних таксонів (від найпростіших до птахів і ссавців). В основі добових ритмів життєдіяльності лежать спадково закріплені ендогенні цикли фізіологічних процесів із періодом, близьким до 24 годин. Циклічні процеси такого роду отримали назву циркадіанних (циркадних) ритмів.
У найбільш «чистому» вигляді циркадні ритми виявляються лише при перебуванні тварин у сталих умовах, тобто без впливу мінливих чинників середовища. Установлено, що ендогенні ритми легко синхронізуються будь-якими зовнішніми датчиками часу (зміни освітленості, температури тощо). Характерна особливість циркадіанних ритмів — деяке неспівпадання їх періоду з повною астрономічною добою. Зокрема, досліди із птахами показали, що при перебуванні денних видів у темряві або в умовах дуже слабкого постійного освітлення добовий цикл активності та рівня метаболізму зберігається тривалий час, але період коливань дещо відрізняється від 24 годин. У результаті цього активність поступово зміщується на інші, ніж у нормальних умовах, години доби. Аналогічні дані отримані й у дослідах з іншими тваринами.
Певний вплив на характер циркадіанних ритмів здійснюють умови освітлення. Збільшення інтенсивності безперервного освітлення викликає у нічних видів зменшення загальної активності, деяке подовження циклу та скорочення його активної частини; при зменшенні освітленості спостерігаються зміщення протилежного характеру. Денні тварини відповідно демонструють зворотні реакції. Ця закономірність отримала назву правила Ашоффа (Ашофф, 1964).
Уважають, що неспівпадання циркадіанного ритму з тривалістю астрономічної доби відкриває можливість зміщення ритмів активності в порядку їх синхронізації з природною зміною умов у кожному конкретному районі у різні періоди року. На цій підставі зовнішні чинники, які виступають як датчики часу, часто називають чинниками-синхронізаторами. Існує й інший погляд, згідно з яким неспівпадання ендогенного ритму з астрономічною добою розглядають як артефакт, зумовлений неприродністю умов експерименту (темрява, незмінність впливаючих чинників тощо).
У дослідах із комахами показано, що за вільного вибору світла та темряви тварини демонструють ритм, що дорівнює 24 годинам. Ці дані трактуються як повний збіг ендогенного ритму з астрономічною добою. Близькі дані отримані у дослідах із деякими птахами: тривалість добового циклу за вільного вибору світла та темряви складала 23,7 години, проте аналогічні досліди із сірим вовчком (Glis glis) виявили циркадіанний характер ритму, що перевищує 24 години. Неспівпадання даних, отриманих для природних умов, і експериментальних даних указують на складний характер явища та не досить повне його вивчення.
У більшості тварин різні фізіологічні та біологічні процеси проявляються сезонно: розмноження, линяння, сплячка та діапауза, міграції тощо. Еволюційно сезонність цих явищ виникла як пристосування до циклічних змін кліматичних умов. Закономірна повторюваність сезонних станів формується у результаті взаємодії вроджених ендогенних сезонних циклів з інформацією про стан зовнішніх умов. Ці взаємодії синхронізують прояв ендогенної програми з періодами сприятливого для даної форми діяльності поєднання чинників середовища і забезпечують адаптацію організму до сезонного стану зовнішніх умов. Фізіологічні механізми формування та регуляції сезонних явищ найкращим чином вивчені у вищих тварин.
Ендогенні біологічні цикли з річною періодичністю, названі циркануальними (цирканними) ритмами, як і циркадні, ґрунтуються на системі вільного відліку часу за принципом біологічного годинника. У природних умовах ця система знаходиться під контролем зовнішніх чинників-синхронізаторів, серед яких у нетропічних тварин головна роль належить фотоперіоду.
Прояв циркануальних ритмів може бути достатньо складним, але у будь-якому разі в них закладений механізм часової програми та контроль із боку природного режиму освітлення. У шовкопряда (Bombyx mori) з яєць, які розвивалися за короткого весняного дня, відроджуються самки, що відкладають яйця, не впадаючи у діапаузу. Яйця, розвиток яких ішов в умовах тривалого літнього дня, дають самок, що відкладають яйця з діапаузою, забезпечуючи таким чином появу весняного покоління.
У штучних умовах, що повністю виключають дію зовнішніх датчиків часу, виявлено, що власний хід циркануального ритму найчастіше буває дещо меншим від астрономічного року. Так, дві славки — садова (Sylvia borin) і чорноголова (S. atricapilla) — у віці 6 тижнів були поміщені в умови постійного фотоперіоду (10 годин світла та 14 — темряви) і перебували у цих умовах відповідно 10 і 8 років. Періоди линьок у цих птахів регулярно повторювалися з періодичністю 9,4—9,7 місяця. Аналогічні досліди з іншими птахами також дали подібні результати. У ховрахів (Citellus lateralis), що протягом трьох років перебували в умовах постійного освітлення та стабільної температури, виявлені циркануальні ритми з періодом 344 доби; у них же виявлена циркануальна періодичність споживання їжі.
Для формування сезонних станів найбільше значення мають гонадотропні гормони (гонадотропіни), що стимулюють функції статевих залоз; тіреотропний гормон, який контролює діяльність щитоподібної залози; адренокортикотропний гормон (АКТГ), що активує продукцію гормонів кори надниркових залоз; пролактин, що бере участь у прямій регуляції розмноження та (у птахів) міграцій. Таким чином, система гіпоталамус — гіпофіз визначає зв’язок фізіологічних сезонних процесів із контролюючою дією фотоперіоду. Фотоперіодичний контроль нейросекреторної активності гіпоталамуса у природних умовах відбувається на базі зорової рецепції. Проте це не обов’язкова умова. Експериментально доведено, що зміни режиму освітлення можуть сприйматися і без участі органів зору на базі світлочутливих структур центральної нервової системи.
Фотоперіодична регуляція сезонних циклів. Механізми фотоперіодичної стимуляції гіпоталамо-гіпофізарної системи достатньо повно вивчені лише на прикладі птахів. За сучасними даними, ефективність дії світлового чинника визначається співвідношенням фотоперіоду та ендогенного циркадіанного ритму гіпоталамічної нейросекреторної системи.
У цілому фотоперіодична регуляція сезонних циклів у хребетних тварин ґрунтується на системі фазових взаємодій їх добових ритмів. При цьому реакція на змінені фотоперіоди може бути різною. Деякі фізіологічні процеси розвиваються негайно після фотостимуляції (наприклад, весняний розвиток гонад). Для інших явищ дата ефективного фотоперіоду слугує початковою «точкою відліку», а сам процес розвивається через певний інтервал часу після такої дії (наприклад, весняний міграційний стан птахів).
Нарешті, у певних випадках ефективна фотоперіодична стимуляція запускає механізм вільного відліку часу, на основі якого у визначені терміни включаються чергові сезонні стани (післяшлюбне линяння, осінній міграційний стан і початок фоторефрактерної фази у птахів). Загальна річна циклічність у цьому випадку представлена цілісною програмою сезонних фізіологічних станів, що змінюють один одного. Роль фотоперіодичної регуляції полягає в координації ендогенного відліку часу із ходом природних періодичних процесів.
У безхребетних тварин (краще досліджені членистоногі) характер фотоперіодичної регуляції дещо інший. Система діє за принципом «пісочного годинника». Для її запуску потрібна специфічна для виду тривалість фотоперіоду. Стимуляція системи сумою коротших періодів світла неможлива.
3.4.3. Водне середовище
На нашій планеті живі організми освоїли чотири основних середовища проживання, які кардинально відрізняються за специфікою умов. Водне середовище було першим, в якому виникло й поширилося життя. У подальшому живі організми оволоділи наземно-повітряним середовищем, утворили й заселили ґрунт. Четвертим специфічним середовищем життя стали самі живі організми, кожний з яких є цілим світом для паразитів або симбіонтів. Вода як середовище існування має ряд специфічних властивостей, таких як висока щільність, істотні перепади тиску, відносно низький вміст кисню, сильне поглинання сонячних променів тощо. Водойми та окремі їх ділянки розрізняються, крім цього, за сольовим режимом, швидкістю горизонтальних переміщень (течій), вмістом зважених часток. Для життя придонних організмів мають значення властивості ґрунту, режим розкладання органічних залишків тощо. Тому, разом з адаптаціями до загальних властивостей водного середовища його мешканці повинні бути пристосовані також до різноманітних окремих умов. Мешканці водного середовища отримали в екології загальну назву гідробіонтів. Вони населяють Світовий океан, континентальні водойми та підземні води. У будь-якій водоймі можна виділити ряд різних за умовами зон.
Екологічні зони Світового океану
В океані та морях розрізняють перш за все дві екологічні зони: товщу води — пелагіаль і дно — бенталь. Залежно від глибини бенталь поділяється на субліторальну зону — область плавного пониження суші до глибини приблизно 200 м, батіальну — область крутого схилу та абісальну зону — область океанічного ложа із середньою глибиною 3—6 км. Ще глибші області бенталі, що відповідають западинам океанічного ложа, називають ультраабісаллю. Частина берега, що заливається водою під час припливів, зветься літораллю. Вища за
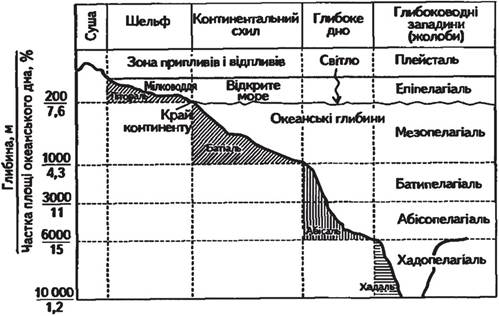
Рис. 3.11. Схема розподілу океанського простору з різними екологічними факторами (Білявський та ін., 2004)
рівень припливів частина берега, що зволожується бризками, отримала назву супраліторалі (рис. 3.11).
Населення дна океану отримало назву бентос. Мешканці субліторалі живуть в умовах відносного невисокого тиску, денного сонячного освітлення, часто досить значних змін температурного режиму. Мешканці абісальних і ультраабі- сальних глибин існують у темряві, за постійної температури та тиску в декілька сотень, а іноді і близько тисячі атмосфер. Тому одна лише вказівка на те, в якій зоні бенталі мешкає той або інший вид організмів, уже говорить про те, які загальні екологічні властивості він повинен мати.
Організми, що мешкають у товщі води (пелагіалі), відносяться до пелагосу. Пелагіаль також поділяють на вертикальні зони, відповідно до того, як поділяються за глибиною зони бенталі: епіпелагіаль, батипелагіаль, абісо- пелагіаль. Нижня межа епіпелагіалі (не глибше 200 м) визначається проникненням сонячного світла в кількості, достатній для фотосинтезу. Фотосинтезуючі рослини глибше цих зон існувати не можуть. У «присмеркових» батіальних і «повних мороку» абісальних глибинах мешкають лише мікроорганізми та тварини.
Основні властивості водного середовища
Щільність води — чинник, що визначає умови пересування водних організмів і тиск на різних глибинах. Для дистильованої води щільність дорівнює 1 г/см3 при +4 °C. Густина природних вод, що містять розчинені солі, може бути більшою — до 1,35 г/см3. Тиск зростає з глибиною приблизно на 1∙105 Па (1 атм) на кожні 10 м.
У зв’язку з різким градієнтом тиску у водоймах гідробіонти в цілому значно більш еврібатні порівняно з суходільними організмами. Деякі види, поширені на різних глибинах, переносять тиск від декількох до сотень атмосфер. Наприклад, голотурії роду Elpidia, черви Priapulus caudatus мешкають від прибережної зони до ультраабісалі. Навіть прісноводі мешканці (інфузорії-туфельки, сувійки, жуки-плавунці тощо) витримують у досліді тиск до 6∙107 Па (600 атм).
Проте багато мешканців морів і океанів відносно стенобатні й приурочені до певних глибин. Стенобатність найчастіше притаманна мілководним і глибоководним видам. Тільки на літоралі мешкають кільчастий черв-піскожил Arenicola, молюски морські блюдечка (Patella). Багато риб, наприклад із групи вудильників, головоногі молюски, ракоподібні, погонофори, морські зірки та інші трапляються лише на великих глибинах за тиску не менше 4—5-107 Па (400—500 атм).
Планктонні організми мають численні адаптації, що підвищують їх плавучість і перешкоджають осіданню на дно. До таких пристосувань відносяться: 1) загальне збільшення відносної поверхні тіла за рахунок зменшення розмірів, сплощеності, подовження, розвитку численних виростів або щетинок, що збільшують тертя об воду; 2) зменшення щільності за рахунок редукції скелета, накопичення в тілі жирів, бульбашок газу тощо.
Щільність і в’язкість води істотно впливають на можливість активного плавання. Тварин, здатних до швидкого плавання та подолання сили течій, об’єднують в екологічну групу нектону (nectos — плаваючий). Представники нектону — риби, кальмари, дельфіни. Швидкий рух у водній товщі можливий лише за наявності обтічної форми тіла та сильно розвиненої мускулатури. Торпедоподібна форма тіла існує в усіх гарних плавців незалежно від їх систематичної належності та способу руху у воді (реактивного, за рахунок вигинання тіла або за допомогою кінцівок).
Кисневий режим. У насиченій киснем воді вміст його не перевищує 10 мл/л, що у 21 раз менше, ніж в атмосфері. Тому умови дихання гідробіонтів значно ускладнені. Верхні шари водної товщі, як правило, багатші киснем, ніж нижні. Із підвищенням температури та солоності води концентрація в ній кисню знижується. У шарах, щільно заселених тваринами та бактеріями, може створюватися різкий дефіцит O2 унаслідок посиленого його споживання. Наприклад, у Світовому океані багаті життям глибини від 50 до 1000 м характеризуються різким погіршенням аерації: вона у 7—10 разів нижча, ніж у поверхневих водах, населених фітопланкто- ном. Серед водних мешканців багато видів, здатних переносити істотні коливання вмісту кисню у воді, аж до майже повної його відсутності (евріоксибіонти — «окси» — кисень, «біонт» — мешканець). До них відносяться, наприклад, прісноводі олігохети Tubifex tubifex, черевоногі молюски Viviparus viviparus. Серед риб дуже слабке насичення води киснем можуть витримувати сазан, лин, карасі. Разом із цим ряд видів — стеноксибіонти — можуть існувати лише за достатньо високого насичення води киснем (райдужна форель, кумжа, гольян, війковий черв Planaria alpina, личинки одноденок, веснянок тощо). Багато видів здатні за нестачі кисню впадати в неактивний стан — аноксибіоз — і таким чином переживати несприятливий період.
Дихання гідробіонтів здійснюється або через поверхню тіла, або через спеціалізовані органи (зябра, легені, трахеї). При цьому покриви можуть служити додатковим органом дихання. Наприклад, в’юн через шкіру споживає у середньому до 63 % кисню. Дихання полегшується також завдяки збільшенню поверхні тіла. Це досягається під час еволюції видів з утворенням у особин різних виростів, сплощень, подовжень, загальним зменшенням розмірів тіла. Деякі види за нестачі кисню активно змінюють величину дихальної поверхні. Черви Tubifex tubifex сильно витягують тіло в довжину, гідри та актинії — подовжують щупальця, голкошкірі — ам- булакральні ніжки. Багато сидячих і малорухливих тварин оновлюють навколо себе воду, або створюючи її направлений потік, або коливальними рухами сприяючи її перемішуванню. Двостулковим молюскам для цієї мети служать війки, що вистилають стінки мантійної порожнини, ракоподібним — черевні чи грудні ніжки. П’явки, личинки комарів-дзвонців (мотиль), багато олігохет розхитують тіло, висунувшись із ґрунту.
У деяких видів трапляється комбінування водного та повітряного дихання: дводишні риби, сифонофори диско- фанти, багато легеневих молюсків, ракоподібні (Gammarus Iacustris) тощо. Вторинноводні тварини зберігають зазвичай атмосферний тип дихання як вигідніший енергетично і тому потребують контактів із повітряним середовищем (ластоногі, китоподібні, водяні жуки, личинки комарів тощо).
Сольовий режим. Підтримання водного балансу гідробіонтів має свою специфіку. Якщо для наземних тварин і рослин найважливіше — забезпечення організму водою в умовах її дефіциту, то для гідробіонтів не менш істотне — збереження певної кількості води в тілі за її надлишку в навколишньому середовищі.
Більшість водних мешканців пойкілосмотичні: осмотичний тиск в їх тілі залежить від солоності навколишньої води. Тому для гідробіонтів основний спосіб підтримувати сольовий баланс — уникати місцеперебування з невідповідною солоністю. Прісноводі форми не можуть існувати в морях, морські — не переносять опріснення. Якщо солоність води змінюється, тварини переміщуються у пошуках сприятливого середовища. Хребетні тварини, вищі раки, комахи, що мешкають у воді, відносяться до гомойосмотичних видів (зберігають постійний осмотичний тиск у тілі незалежно від концентрації солей у воді).
У прісноводих видів рідини тіла гіпертонічні відносно навколишнього водного середовища. Їм загрожує зайве обводнення, якщо не перешкоджати надходженню або не викидати надлишок води з тіла. У найпростіших це досягається роботою видільних вакуолей, у багатоклітинних — виділенням води через відповідну систему. Деякі інфузорії, наприклад, кожні 2,0—2,5 хв виділяють кількість води, яка дорівнює масі їхнього тіла.
Якщо ж вода гіпертонічна відносно рідин тіла гідробіонтів, їм загрожує зневоднення в результаті осмотичних втрат. Захист від зневоднення досягається підвищенням концентрації солей у тілі гідробіонтів. Зневодненню перешкоджають також і непроникні для води покриви гомойосмотичних організмів (ссавців, риб, вищих ракоподібних, водних комах).
За дефіциту води в тілі внаслідок зростання солоності багато пойкілосмотичних видів переходить до неактивного стану (анабіозу). Це властиво видам, що мешкають у калюжах морської води та на літоралі (коловерткам, джгутиковим, інфузоріям, деяким рачкам, чорноморським поліхетам Nereis divesicolor тощо). Сольовий анабіоз — спосіб переживати несприятливі умови внаслідок змінної солоності води. Істинно еврігалінних видів, здатних в активному стані мешкати як у прісній, так і в солоній воді, серед водних мешканців не так багато. В основному це види, що населяють естуарії річок, лимани та інші солонуватоводні водойми.
Температурний режим водойм стабільніший, ніж на суходолі. Це пов’язано з фізичними властивостями води, перш за все із високою її питомою теплоємністю, завдяки якій отримання або віддача значної кількості тепла не призводить до різких змін її температури. Амплітуда річних коливань температури у поверхневих шарах океану не більше 10—15 °C, у континентальних водоймах — 30—35 °C. Глибші шари води відрізняються постійністю температури. В екваторіальних водах середньорічна температура поверхневих шарів +26...+27 °С, у полярних — близько 0 °C і нижче. У гарячих наземних джерелах температура води може досягати + 100 °C, а в підводних гейзерах за високого тиску на дні океану навіть зареєстрована температура +380 °C.
Таким чином, у водоймах існує досить значна різноманітність температурних умов. Між верхніми шарами води з вираженими в них сезонними коливаннями температури та нижніми, де тепловий режим постійний, існує зона температурного стрибка, або термоклину. Термоклин різкіше виражений у теплих морях, де сильніший перепад температури поверхневих і глибинних вод.
У зв’язку зі стійкішим температурним режимом води серед гідробіонтів значно більшою мірою, ніж серед населення суходолу, поширена стенотермність. Еврітермні види трапляються в основному в невеликих континентальних водоймах і на літоралі морів високих і помірних широт, де зафіксовані істотні добові та сезонні коливання температури.
Світловий режим. Світла у воді значно менше, ніж у повітрі. Частина падаючих на поверхню водойми променів відбивається у повітряне середовище. Віддзеркалення тим сильніше, чим нижча висота сонця над горизонтом, тому день під водою коротший, ніж на суходолі. Наприклад, літній день поблизу острова Мадейра на глибині 30 м — 5 годин, а на глибині 40 м — лише 15 хвилин. Сутінки, що згущуються в океані з глибиною, мають спочатку зелений, потім блакитний, синій і синьо-фіолетовий колір, змінюючись нарешті постійною темрявою. Відповідно змінюють один одного з глибиною зелені, бурі та червоні водорості, спеціалізовані на поглинанні світла різної довжини хвилі.
Забарвлення тварин міняється з глибиною так само закономірно. Найяскравіше і найрізноманітніше забарвлені мешканці літоральної та субліторальної зон. Багато глибинних організмів, подібно до печерних, не мають пігментів. У смерковій зоні найбільш поширене червоне забарвлення, яке є додатковим до синьо-фіолетового світла на цих глибинах. Це дозволяє мешканцям ховатися від ворогів, оскільки їхній червоний колір у синьо-фіолетових променях сприймається як чорний. Червоне забарвлення характерне для таких тварин смеркової зони, як морський окунь, червоний корал, різні ракоподібні тощо.
У деяких видів, що мешкають поблизу поверхні водойм, очі розділяються на дві частини з різною здатністю до заломлення променів світла. Одна половина ока (верхня) бачить у повітрі, нижня — у воді. Така «чотириокість» характерна для жуків-вертячок, американської рибки Anableps tetrophthalmus, одного із тропічних видів морських собачок Dialommusfuscus (рис. 3.12).

Рис. 3.12. Риби, очі яких пристосовані бачити як у воді, так і в повітрі (Нікольський, 1974): а— чотириока риба Anableps Ietrophthalmus; б — чотириока риба морська собачка Dialomnus fuscus
Поглинання світла тим сильніше, чим менша прозорість води, яка залежить від кількості завислих у ній часток. Прозорість характеризують граничною глибиною, на якій ще видно білий диск діаметром близько 20 см (диск Секкі). Найпрозоріші води у Саргасовому морі: диск видно до глибини 66,5 м. У Тихому океані диск Секкі видно до 59 м, в Індійському — до 50, у невеликих морях — до 5—15 м. Прозорість річок у середньому 1,0—1,5 м, а в найкаламут- ніших річках, наприклад у середньоазіатських Амудар’ї та Сирдар’ї, — лише декілька сантиметрів. Саме тому межа зони фотосинтезу сильно варіює в різних водоймах. У найчистіших водах еуфотична зона (зона фотосинтезу) сягає глибин понад 200 м, дисфотична (смеркова зона) займає глибини до 1000—1500 м. Глибше, до афотичної зони, сонячне світло не проникає зовсім.
У темних глибинах океану як джерело зорової інформації організми використовують світло, що продукують живі істоти. Світіння живого організму отримало назву біолюмі- несценція. Види, що світяться, є майже в усіх класах водних тварин від найпростіших до риб, а також серед бактерій, нижчих рослин і грибів. Хімія біолюмінесценції нині досить добре вивчена. Реакції, що використовуються для генерації світла, різноманітні. Але в усіх випадках це окиснення складних органічних сполук люциферинів за допомогою білкових каталізаторів люцифераз. Під час реакції надлишок енергії збудженої молекули люциферину виділяється у вигляді квантів світла.
Світіння може і не відігравати особливої екологічної ролі у житті виду, а бути побічним результатом життєдіяльності клітин, як, наприклад, у бактерій або нижчих рослин. Екологічну сутність воно отримує тільки у тварин, що мають достатньо розвинену нервову систему та органи зору. У багатьох видів органи світіння набувають дуже складної будови з системою відбивачів і лінз, що посилюють випромінювання (рис. 3.13). Низка риб і головоногих молюсків, не здатних генерувати світло, використовує симбіотичних бактерій, що розмножуються у спеціальних органах цих тварин.

Рис. 3.13. Органи світіння водних тварин (Нікольський, 1974): 1 — розподіл органів світіння на тілі глибоководної риби Lampanyctes; 2 — світний орган глибоководної риби Argyropelecus affinis: а — пігмент, б — рефлектор, в — світне тіло, г — лінза
Біолюмінесценція має в житті тварин в основному сигнальне значення. Світлові сигнали можуть слугувати для орієнтації у зграї, залучення особин різної статі, підманювання жертв, маскування або втечі. Наприклад, спалах світла може бути захистом від хижака, засліплюючи його або дезорієнтуючи. Глибоководні каракатиці, рятуючись від ворога, випускають хмару секрету, що світиться, тоді як види, що мешкають в освітлених водах, використовують для цієї мети темну рідину. У деяких донних багатощетинкових чер- вів-поліхет органи, що світяться, розвиваються до періоду дозрівання статевих продуктів, причому світяться яскравіше у самок, а очі краще розвинені у самців. У хижих глибоководних риб із ряду вудильникових перший промінь спинного плавця зміщений до верхньої щелепи і перетворений на гнучке «вудилище», що несе на кінці червоподібну приманку — залозу, заповнену слизом із бактеріями, що світяться. Регулюючи приплив крові до залози і, отже, постачання бактерій киснем, риба може довільно викликати світіння «приманки», а імітуючи рухи черв’яка — підманювати здобич.
У наземній обстановці біолюмінесценція розвинена лише у небагатьох видів, найсильніше — у жуків із родини світляків, які використовують світлову сигналізацію для приманювання особин іншої статі у присмеркові або нічні години.
Способи орієнтації тварин у водному середовищі
Життя у постійних сутінках або мороці сильно обмежує можливості зорової орієнтації гідробіонтів. У зв’язку зі швидким згасанням світлових променів у воді тварини навіть із добре розвиненими органами зору орієнтуються за їх допомогою лише на близькій відстані.
Звук розповсюджується у воді швидше, ніж у повітрі. Орієнтація на звук розвинена у гідробіонтів у цілому краще, ніж зорова. Ряд видів уловлює навіть коливання дуже низької частоти (інфразвуки), що виникають при зміні ритму хвиль, і завчасно опускається перед штормом із поверхневих шарів води до глибших (наприклад, медузи). Багато мешканців водойм (ссавці, риби, молюски, ракоподібні) самі видають звуки. Ракоподібні здійснюють це тертям одне об одного різних частин тіла; риби — за допомогою плавального міхура, глоткових зубів, щелеп, променів грудних плавників, іншими способами. Звукова сигналізація служить найчастіше для внутрішньовидових взаємин, наприклад для орієнтації у зграї, залучення особин іншої статі й особливо розвинена у мешканців каламутних вод і великих глибин.
Ряд гідробіонтів розшукує їжу та орієнтується за допомогою ехолокації — сприйняття відбитих звукових хвиль (китоподібні). Багато хто сприймає відбиті електричні імпульси, створюючи при плаванні електричні розряди різної частоти. Відомо близько 300 видів риб, здатних генерувати електрику та використовувати її для орієнтації і сигналізації. Прісновода рибка водяний слон (Mormyrus kannume) посилає до 30 імпульсів за секунду, виявляючи безхребетних, яких вона знаходить у рідкому мулі без допомоги зору. Частота розрядів у деяких морських риб доходить до 2000 імпульсів за секунду. Ряд риб використовує електричні поля також для захисту та нападу (електричний скат, електричний вугор та інші).
Для орієнтації в глибині служить сприйняття гідростатичного тиску за допомогою статоцистів, газових камер, інших спеціалізованих органів.
Найдавніший спосіб орієнтації, властивий всім водним тваринам, — сприйняття хімізму середовища. Хеморецептори багатьох гідробіонтів мають надзвичайну чутливість. У тисячокілометрових міграціях, характерних для багатьох видів риб, вони орієнтуються в основному за запахом, із вражаючою точністю знаходячи місця нерестовищ або нагулу. Експериментально доведено, наприклад, що лососі, штучно позбавлені нюху, не знаходять гирла своєї річки, повертаючись на нерест, але ніколи не помиляються, якщо можуть сприймати запахи. Надзвичайно тонкий нюх у риб, що здійснюють особливо далекі міграції.
3.4.4. Наземно-повітряне середовище життя
Наземно-повітряне середовище — найскладніше за екологічними умовами. Життя на суходолі вимагало таких пристосувань, які виявилися можливими лише за достатньо високого рівня організації як рослин, так і тварин.
Повітря як екологічний чинник для наземних організмів. Низька щільність повітря визначає його малу підйомну силу та незначну силу опору. Мешканці повітряного середовища повинні мати власну опорну систему, що підтримує тіло: рослини — різноманітні механічні тканини, тварини — твердий або, значно рідше, гідростатичний скелет. Крім того, всі мешканці повітряного середовища тісно пов’язані з поверхнею землі, яка служить їм для прикріплення та опори. Життя у завислому стані у повітрі неможливе.
Безліч мікроорганізмів і тварин, спори, насіння, плоди та пилок рослин регулярно присутні у повітрі й розносяться повітряними течіями, багато тварин здатні до активного польоту, проте в усіх цих видів основна функція їх життєвого циклу — розмноження — здійснюється на поверхні ґрунту. Для більшості з них перебування у повітрі пов’язане тільки з розселенням або пошуком здобичі.
Мала щільність повітря обумовлює низьку опірність пересуванню. Тому багато наземних тварин використовували у ході еволюції екологічні вигоди цієї властивості середовища, набувши здатність до польоту. До активного польоту здатні близько 75 % видів наземних тварин (переважно комахи та птахи, але трапляються «літуни» і серед ссавців і рептилій). Літають наземні тварини в основному за допомогою мускульних зусиль, але деякі можуть і планерувати за рахунок повітряних течій.
У багатьох видів розвинена анемохорія — розселення за допомогою повітряних потоків. Анемохорія характерна для спор, насіння та плодів рослин, цист найпростіших, дрібних комах, павуків, ракоподібних. Організми, що пасивно переносяться потоками повітря, отримали у сукупності назву аеропланктону за аналогією з планктонними мешканцями водного середовища. Спеціальні адаптації для пасивного польоту: дрібні розміри тіла, збільшення площі його поверхні за рахунок виростів, сильного розчленування тіла, великої відносної поверхні крил, використання павутини тощо.
Мала щільність повітря обумовлює також порівняно низький тиск на суходолі. У нормі він дорівнює 760 мм рт. ст. Зі збільшенням висоти над рівнем моря тиск зменшується. На висоті 5800 м він дорівнює лише половині нормального. Таким чином, низький тиск може обмежувати розповсюдження видів у горах. Для більшості хребетних верхня межа життя знаходиться на висоті близько 6000 м н. р. м. Зниження тиску спричиняє зменшення забезпеченості киснем і зневоднення тварин за рахунок збільшення частоти дихання. Приблизно такі ж межі просування в горах у вищих рослин. Дещо витриваліші членистоногі (ногохвістки, кліщі, павуки), які можуть траплятися на льодовиках, вище за межу поширення рослинності. У цілому всі наземні організми набагато більш стенобатні, ніж водні, оскільки звичайні коливання тиску навколишнього середовища становлять долі атмосфери і навіть для птахів, що піднімаються на велику висоту, не перевищують 1/3 нормального.
Газовий склад повітря. Окрім фізичних властивостей повітряного середовища, для існування наземних організмів надзвичайно важливіші його хімічні особливості. Газовий склад повітря у приземному шарі атмосфери досить однорідний відносно вмісту головних компонентів (азот — 78,1 %, кисень — 21,0, аргон — 0,9, вуглекислий газ — 0,035 % за об’ємом) завдяки високій дифузійній здатності газів і постійному перемішуванню конвекційними та вітровими потоками.
Високий вміст кисню сприяв підвищенню обміну речовин у наземних організмів порівняно з первинноводними. Саме у наземному середовищі, на базі високої ефективності окислювальних процесів в організмі, виникла гомойотермія тварин.
Вміст вуглекислого газу може змінюватися в окремих ділянках приземного шару повітря в досить значних межах. Наприклад, за відсутності вітру в центрі великих міст концентрація його збільшується в десятки разів. Закономірні добові зміни вмісту вуглекислоти в приземних шарах пов’язані з ритмом фотосинтезу рослин. Сезонні зміни обумовлені змінами інтенсивності дихання живих організмів, переважно мікроскопічного населення ґрунтів. Підвищене насичення повітря вуглекислим газом спостерігається у зонах вулканічної активності, поблизу термальних джерел та інших підземних виходів цього газу.
У природі основне джерело вуглекислоти — так зване ґрунтове дихання. Ґрунтові мікроорганізми та тварини дихають дуже інтенсивно. Вуглекислий газ дифундує з ґрунту до атмосфери, особливо активно під час дощу. Багато його виділяють ґрунти помірно вологі, такі, що добре прогріваються, багаті органічними залишками. Наприклад, ґрунт букового лісу виділяє CO2 від 15 до 22 кг/га за годину, а збіднений піщаний — лише 2 кг/га за годину.
Азот повітря для більшості мешканців наземного середовища — інертний газ, але низка прокаріотичних організмів (бульбочкові бактерії, азотобактер, клостридії, ціанобактерії тощо) мають здатність асимілювати його та залучати до біологічного кругообігу.
3.4.5. Ґрунт і рельєф
Едафічні чинники середовища. Властивості ґрунту та рельєф місцевості впливають на умови життя наземних організмів, у першу чергу рослин. Властивості земної поверхні, що здійснюють вплив на її мешканців, об’єднують назвою едафічні чинники середовища (від грецького «едафон» — підстава, ґрунт). Рельєф місцевості та характер ґрунту впливають на специфіку пересування тварин. Наприклад, копитні, страуси, дрохви, що живуть на відкритих просторах, потребують твердого ґрунту для посилення відштовхування під час швидкого бігу. У ящірок, що мешкають на сипких пісках, пальці облямовані бахромою з рогових лусочок, яка збільшує поверхню опори. Для наземних мешканців, що риють нори, щільні ґрунти несприятливі. Отже характер ґрунту в ряді випадків впливає на розподіл наземних тварин, що риють нори, зариваються в ґрунт для порятунку від спеки або хижаків, відкладають у ґрунті яйця.
Ґрунт як місце існування. Особливості ґрунту. Ґрунт — пухкий тонкий поверхневий шар суходолу, що контактує з повітряним середовищем. Незважаючи на незначну товщину, ця оболонка Землі відіграє найважливішу роль у розповсюдженні життя. Ґрунт — не просто тверде тіло, як більшість порід літосфери, а складна трифазна система, в якій тверді частки оточені повітрям і водою. Він пронизаний порожнинами, заповненими сумішшю газів і водними розчинами, і тому в ньому складаються надзвичайно різноманітні умови, сприятливі для життя безлічі мікро- та макроорганізмів. У ґрунті згладжені температурні коливання порівняно з приземним шаром повітря, а наявність ґрунтових вод і проникнення опадів створюють запаси вологи та забезпечують режим вологості, проміжний між водним і наземним середовищем. У ґрунті концентруються запаси органічних і мінеральних речовин, що постачаються відмираючою рослинністю та трупами тварин. Усе це визначає велику насиченість ґрунту життям.
У середньому у 1 м2 ґрунтового шару міститься понад 100 млрд клітин найпростіших, мільйони коловерток і тихоходів, десятки мільйонів нематод, десятки й сотні тисяч кліщів і колембол, тисячі інших членистоногих, десятки тисяч енхитреїд, десятки й сотні дощових червів, молюсків та інших безхребетних. Крім того, 1 см2 ґрунту містить десятки й сотні мільйонів бактерій, мікроскопічних грибів, актиноміцетів та інших мікроорганізмів.
Неоднорідність умов у ґрунті найчіткіше виявляється у вертикальному напрямі. Із глибиною різко змінюється ряд найважливіших екологічних чинників, що впливають на життя мешканців ґрунту. Перш за все це відноситься до структури ґрунту. У ній виділяють три основних горизонти, що розрізняються за морфологічними та хімічними властивостями:
1) верхній, перегнійно-акумулятивний горизонт А, у якому накопичується й перетворюється органічна речовина і з якого промивними водами частина сполук виноситься донизу;
2) ілювіальний горизонт В, де осідають і перетворюються вимиті речовини;
3) материнську породу, або горизонт С, матеріал якої перетворюється на ґрунт.
У межах кожного горизонту виділяються дрібніші шари, що також істотно відрізняються за властивостями. Наприклад, у зоні помірного клімату під хвойними або змішаними лісами горизонт А складається з підстилки (A0) — шару пухкого скупчення рослинних залишків, темнозабарвленого гумусового шару (A1), в якому частки органічного походження перемішані з мінеральними, і підзолистого шару (A2), попелясто-сірого за кольором, в якому переважають сполуки кремнію, а всі розчинні речовини вимиті у глиб ґрунтового профілю (елювіальний шар). Як структура, так і хімізм цих шарів дуже різні, тому коріння рослин і мешканці ґрунту, переміщаючись лише на декілька сантиметрів угору або донизу, потрапляють в інші умови.
Розміри порожнин між частками ґрунту, придатних для проживання в них тварин, зазвичай швидко зменшуються з глибиною. Наприклад, у лучних ґрунтах середній діаметр порожнин на глибині 0—1 см сягає 3 мм, 1—2 см — 2 мм, а на глибині 2—3 см — лише 1 мм; глибше ґрунтові пори ще дрібніші. Щільність ґрунту також змінюється з глибиною. Найпухкіші шари ті, що містять органічну речовину. Порожнистість цих шарів визначається тим, що органічні речовини склеюють мінеральні частки у крупніші агрегати, об’єм порожнин між якими збільшується. Найщільніший зазвичай ілювіальний горизонт В, зцементований вимитими до нього колоїдними частками.
Волога у ґрунті наявна у різних станах: 1) зв’язана (гігроскопічна та плівкова) — міцно утримується поверхнею ґрунтових часток; 2) капілярна — займає дрібні пори і може пересуватися по них у різних напрямах; 3) гравітаційна — заповнює крупніші порожнини і поволі просочується вниз під впливом сили тяжіння; 4) пароподібна — міститься у ґрунтовому повітрі.
Вміст води неоднаковий у різних ґрунтах і в різний час. Якщо дуже багато гравітаційної вологи, то режим ґрунту близький до режиму водойм. У сухому ґрунті залишається тільки зв’язана вода, а умови наближаються до наземних. Проте навіть у найсухіших ґрунтах повітря завжди вологіше, ніж наземне, тому мешканці ґрунту значно менш чутливі до загрози висихання, ніж ті, що мешкають на поверхні.
Склад ґрунтового повітря досить мінливий. Із глибиною в ньому значно падає вміст кисню й зростає концентрація вуглекислого газу. У зв’язку з присутністю у ґрунті органічних речовин, що розкладаються, у ґрунтовому повітрі може бути висока концентрація таких токсичних газів, як аміак, сірководень, метан. При затопленні ґрунту або інтенсивному гнитті рослинних залишків місцями можуть виникати повністю анаеробні умови.
Коливання температури значні лише на поверхні ґрунту. Тут вони можуть бути навіть сильніші, ніж у приземному шарі повітря. Проте з кожним сантиметром углиб добові та сезонні температурні коливання стають все меншими і на глибині 1,0—1,5 м практично вже не простежуються.
Усі ці особливості призводять до того, що навіть за наявності істотної неоднорідності екологічних умов у ґрунті він виступає як достатньо стабільне середовище, особливо для рухливих організмів. Тренд градієнтів температури та вологості у ґрунтовому профілі дозволяє ґрунтовим тваринам шляхом незначних переміщень забезпечувати собі відповідні екологічні умови.
Мешканці ґрунту. Неоднорідність ґрунту призводить до того, що для організмів, різних за розмірами, він виступає як різне середовище. Для мікроорганізмів особливе значення має величезна сумарна поверхня ґрунтових часток, оскільки на них адсорбується переважна частина мікробного населення. Складність ґрунтового середовища створює велике різноманіття умов для дуже різних функціональних груп: аеробів і анаеробів, споживачів органічних і мінеральних сполук. Для розподілу мікроорганізмів у ґрунті характерні дрібні осередки, оскільки навіть упродовж декількох міліметрів параметри ґрунтового середовища можуть змінюватись.
Для дрібних ґрунтових тварин, яких об’єднують під назвою мікрофауна (найпростіші, коловертки, тихоходи, нематоди тощо), ґрунт — система мікроводойм. По суті, це водні організми. Вони живуть у ґрунтових порах, заповнених гравітаційною або капілярною водою, а частину життя можуть, як і мікроорганізми, знаходитись в адсорбованому вигляді на поверхні часток у тонких прошарках плівкової вологи (рис. 3.14).
Багато представників таких видів мешкає й у звичайних водоймах. Проте ґрунтові форми набагато дрібніші, ніж прісноводні, і, крім цього, відрізняються здатністю тривалий час знаходитись в інцистованому стані, переживаючи несприятливі періоди. Порівняння розмірів представників мікрофауни ґрунту та вільноживучих тварин свідчать про те, що прісноводі амеби мають розміри 50—100 мкм, ґрунтові — лише

Рис. 3.14. Мікрофауна ґрунту (Чернова, Билова, 2004): 1—4 — джгутикові, 5—8 — голі амеби, 9—10 — черепашкові амеби, 11—13 — інфузорії, 14—16 — круглі черви, 17—18 — коловертки, 19—20 — тихоходи
10—15. Особливо дрібні представники джгутикових (нерідко 2—5 мкм). Ґрунтові інфузорії також мають карликові розміри і до того ж можуть сильно міняти форму тіла.
Для дихаючих повітрям дещо крупніших тварин ґрунт є немовби системою дрібних печер. Таких тварин об’єднують під назвою мезофауна. Розміри представників мезофауни ґрунтів — від десятих часток до 2—3 мм. До цієї групи відносяться в основному членистоногі: численні групи кліщів, первиннобезкрилі комахи (колемболи, протури, двохвістки), дрібні види крилатих комах, багатоніжки симфіли тощо (рис. 3.15).
У них немає спеціальних пристосувань до риття, вони повзають по стінках ґрунтових порожнин за допомогою кінцівок або червоподібно звиваючись. Насичене водяною парою ґрунтове повітря дозволяє цим тваринам дихати через покриви тіла. Такі тварини чутливі до висихання, а основний засіб порятунку від коливання вологості повітря для них — пересування у глиб ґрунтового профілю. Ра-

Рис. 3.15. Мезофауна ґрунту (Чернова, Былова, 2001): 1 — псев- доскорпіон, 2 — гамазовий кліщ, 3—4 — панцирні кліщі, 5 —багатоніжка, 6 — личинка комара-хірономіди, 7 — жук із родини Ptiliidae, 8—9 — колемболи
зом із цим, можливість міграцій по ґрунтових порожнинах углиб обмежується швидким зменшенням діаметра пор, тому пересування по порожнинах ґрунту доступні тільки найдрібнішим видам. Крупніші представники мезофауни мають деякі пристосування, що дозволяють переносити тимчасове зниження вологості ґрунтового повітря. Це захисні лусочки на тілі, часткова непроникність покривів, суцільний товстостінний панцир з епікутикулою у поєднанні з примітивною трахейною системою, що забезпечує дихання.
Періоди затоплення ґрунту водою представники мезофауни переживають у бульбашках повітря. Повітря затримується навколо тіла тварин завдяки їх покривам, що не змочуються, забезпечених до того ж волосками або лусочками. Бульбашка повітря служить для дрібної тварини своєрідною «фізичною зяброю». Дихання відбувається за рахунок кисню, що дифундує в повітряний прошарок із навколишньої води. Представники мікро- і мезофауни здатні переносити зимове промерзання ґрунту, оскільки більшість видів не може перейти донизу із шарів, що піддаються дії негативних температур.
Більших ґрунтових тварин (із розмірами тіла 2—50 мм) називають представниками макрофауни. Це личинки комах, багатоніжки, енхетреїди, дощові черви тощо (рис. 3.16). Для них ґрунт — щільне середовище, що чинить значний ме-
Рис. 3.16. Макрофауна ґрунту (Чернова, Билова, 2001): 1 — дощовий черв, 2 — мокриця, 3 — губонога багатоніжка, 4 — двопарнонога багатоніжка, 5 — личинка туруна, 6 — личинка жука-ковалика, 7 — вовчок, 8 — личинка пластинчатовусого жука
ханічний опір під час руху. Ці відносно великі форми пересуваються у ґрунті або розширюючи природні порожнини шляхом розсування ґрунтових часток, або риючи нові ходи. Обидва способи пересування накладають відбиток на зовнішню будову тварин. Можливість рухатися по тонких порожнинах, майже не вдаючись до риття, властива тільки видам, які мають тіло з малим поперечним перетином, що здатне сильно згинатися у звивистих ходах (багатоніжки-кістянки та геофіли).
Розсовуючи частки ґрунту за рахунок тиску стінок тіла, пересуваються дощові черви, личинки комарів-довгоніжок та інші. При цьому позаду тварини залишається відкритий хід, що загрожує збільшенням випаровування та переслідуванням хижаків. У багатьох видів розвинені пристосування для екологічно вигіднішого типу пересування у ґрунті — риття із закупорюванням за собою ходу. Риття здійснюється розпушуванням і відгортанням ґрунтових часток. Личинки різних комах використовують для цього передній кінець голови, мандибули та передні кінцівки, розширені й укріплені товстим шаром хітину, шпильками та виростами. На задньому кінці тіла розвиваються пристосування для міцної фіксації — підпірки, що висуваються, зубці, гачки. Закриваючи за собою хід, тварини — мешканці ґрунту — постійно знаходяться у замкненій камері, насиченій випаровуваннями власного тіла. Газообмін більшості видів цієї екологічної групи відбувається за допомогою спеціалізованих органів дихання, але доповнюється газообміном через покриви. Можливе навіть лише шкірне дихання, наприклад у дощових червів і енхетреїд. Рийні тварини можуть залишати шари, де виникає несприятлива ситуація. У посуху та до зими вони концентруються у глибших шарах, зазвичай у декількох десятках сантиметрів від поверхні.
Мегафауна ґрунтів — великі землериї, в основному із групи ссавців. Ряд видів проводить у ґрунті все життя (сліпаки, сліпачки, цокори, кроти Євразії, золотокроти Африки, сумчасті кроти Австралії). Вони прокладають у ґрунті цілі системи ходів і нір. Зовнішній вигляд і анатомічні особливості цих тварин відображають їх пристосованість до рийного підземного способу життя. У них недорозвинені очі, компактне, валькувате тіло з короткою шиєю, коротке густе хутро, сильні копальні кінцівки з міцними кігтями. Сліпаки та сліпачки розпушують землю різцями. До мегафауни ґрунту слід віднести також великих олігохет, особливо представників родини Megascolecidae, що мешкають у тропіках Південної півкулі. Найбільший із них австралійський Megascolides australis досягає в довжину 2,5 і навіть 3 м.
Крім постійних мешканців ґрунту, серед великих тварин можна виділити екологічну групу мешканців нір (ховрахи, байбаки, тушканчики, кролі, борсуки тощо). Вони живляться на поверхні, але розмножуються, зимують, відпочивають, рятуються від небезпеки у ґрунті. Ряд інших тварин використовує їх нори, знаходячи в них сприятливий мікроклімат і укриття від ворогів. Нірники мають риси будови, властиві наземним тваринам, але характеризуються і рядом пристосувань, пов’язаних із рийним способом життя. Наприклад, у борсуків довгі кігті та сильна мускулатура передніх кінцівок, вузька голова, невеликі вушні раковини. У кролів порівняно із зайцями, що не риють нір, помітно вкорочені вуха та задні кінцівки, міцніший череп, сильніше розвинені кістки та мускулатура передпліччя.
За цілою низкою екологічних особливостей ґрунт — середовище, проміжне між водним і наземним. Із водним середовищем ґрунт зближують його температурний режим, знижений уміст кисню у ґрунтовому повітрі, насиченість його водяною парою та наявність води в інших формах, присутність солей і органічних речовин у ґрунтових розчинах, можливість рухатися в ньому у трьох вимірах. Із повітряним середовищем ґрунт зближують наявність ґрунтового повітря, загроза висушування у верхніх горизонтах, досить різкі зміни температурного режиму поверхневих шарів.
3.4.6. Погодні та кліматичні особливості наземно-повітряного середовища
Погодні особливості. Умови життя в наземно-повітряному середовищі ускладнюються, крім того, погодними змінами. Погода — безперервно змінний стан атмосфери поблизу земної поверхні до висоти приблизно 20 км (межа тропосфери). Мінливість погоди виявляється у постійному варіюванні поєднання таких чинників середовища, як температура та вологість повітря, хмарність, опади, сила та напрям вітру. Для погодних змін разом із закономірним чергуванням їх у річному циклі характерні неперіодичні коливання, що істотно ускладнює умови існування наземних організмів. На життя водних мешканців погода впливає значно меншою мірою, має суттєвий вплив лише на мешканців поверхневих шарів води.
Клімат місцевості. Багаторічний режим погоди характеризує клімат місцевості. До поняття клімату входять не тільки середні значення метеорологічних явищ, а і їх річний і добовий хід, відхилення від нього та повторюваність. Клімат визначається географічними умовами району.
Для більшості наземних організмів, особливо дрібних, важливий не стільки клімат регіону, скільки умови їх безпосереднього мешкання. Дуже часто місцеві елементи середовища (рельєф, експозиція, рослинність) так змінюють у конкретній ділянці режим температури, вологості, освітленості, руху повітря, що він значно відрізняється від кліматичних умов місцевості. Такі локальні модифікації клімату, що складаються у приземному шарі повітря, називають мікрокліматом. У кожній зоні мікроклімат значно відрізняється. Можна виділити мікроклімат будь-яких ділянок площі, об’єктів. Наприклад, особливий режим утворюється у віночках квіток, який використовують комахи, що мешкають там. Широко відомі відмінності температури, вологості повітря та сили вітру на відкритому просторі й у лісі, у трав’яному ярусі та над оголеними ділянками ґрунту, на схилах північної та південної експозицій. Особливий стійкий мікроклімат виникає в норах, гніздах, дуплах, печерах та інших закритих місцях.
Опади. Крім водозабезпечення та створення запасів вологи, опади можуть мати й іншу екологічну роль. Сильні зливові дощі або град здійснюють іноді механічний вплив на рослини або тварин, затоплюють нори рийних тварин тощо. Особливо різноманітна екологічна роль снігового покриву. Добові коливання температур відчутні у товщі снігу лише до 25 см, глибше температура майже не змінюється. При морозах у —20...—30 °C під шаром снігу товщиною 30—40 см температура не набагато нижча за нуль. Дрібні наземні звірі й узимку ведуть активний спосіб життя, прокладаючи під снігом і в його товщі цілі галереї ходів. Для ряду видів, що живляться підсніжною рослинністю, характерне навіть зимове розмноження, яке відзначається, наприклад, у лемінгів, лісової миші, мишака жовтогорлого, ряду видів нориць, водяного щура та інших ссавців.
Великим тваринам зимовий сніговий покрив заважає добувати корм. Багато копитних (північні олені, кабани, вівцебики) живляться взимку переважно підсніжною рослинністю, і глибокий сніговий покрив, а особливо тверда кірка на його поверхні, що виникає в ожеледь, прирікають їх на нестачу кормів. Глибина снігового покриву може істотно обмежувати розповсюдження видів. Наприклад, справжні олені не проникають на північ у ті райони, де товщина снігу взимку більша 40—50 см.
Білизна снігового покриву демаскує темних тварин. У виникненні сезонної зміни забарвлення у білої тундрової куріпки, зайця-біляка, горностая, ласки, песця, мабуть, велику роль зіграв відбір на маскування під колір фону.
Магнітне поле Землі. Вважається, що основне позитивне значення магнітного поля Землі полягає в тому, що воно, як парасолька, надійно захищає все живе на Землі від жорсткого сонячного та космічного випромінювання. І оскільки живі істоти і людина в тому числі, на 70—80 % складаються з води, то його коливання впливає на електропровідність клітинної протоплазми та тканин і, як наслідок, — на всі біофізичні та біохімічні процеси. Багатьма дослідниками на різних живих об’єктах показано, що екранування природного магнітного поля призводить до їх загибелі.
Власна пульсація магнітного поля Землі (так звана частота Шумана) тривалий час була постійною величиною і дорівнювала 7,83 Гц. Ця частота була настільки стабільною, що слугувала еталоном для навігаційного обладнання літаків, кораблів тощо. На цю ж частоту налаштований і альфа-ритм головного мозку людини. Але в останні десятиріччя вона почала зростати: нині вже наближається до 14 Гц, що відповідає частоті бета-ритму головного мозку, яка пов’язана не зі станом спокою, а зі станом активності людини. Таким чином, ритм «серцебиття» нашої планети невпинно зростає. Цікаво, що разом із тим напруга магнітного поля падає. За останні 22 роки вона зменшилась у середньому на 1,7 %, а в деяких регіонах Атлантичного океану — навіть на 10 %. У США (НАСА) і Німеччині (інститут М. Планка) проводили тривалі експерименти, в результаті яких встановлено, що хвилі Шумана необхідні для синхронізації біологічних ритмів і нормального існування всього живого на Землі. Таким чином, власне магнітне поле Землі — необхідний, життєво важливий фактор існування живих істот на нашій планеті.
Сьогодні вже відомо, що люди, які відчувають великі навантаження та стрес, потребують шуманівських хвиль. Гостро відчувають їх відсутність літні та вегетативно чутливі люди, а також хронічні хворі. Це може призводити до головного болю, втрати орієнтації, нудоти, запаморочення тощо.
Нині електромагнітний фон планети в результаті діяльності людини зазнав істотних змін і забруднений настільки, що хвилі Шумана губляться серед найрізноманітніших електромагнітних випромінювань. Одні з найпоширеніших і найвпливовіших — електромагнітні коливання, викликані промисловими струмами з частотою 50 Гц, а також гігагер- цові частоти стільникового зв’язку. До цього слід додати й те, що природне магнітне поле екранується, насамперед, у будівлях, що мають залізобетонні конструкції та металеві дахові покриття. Отже, сучасна людина загалом не зазнає оптимізувального впливу цього фактора, і тому потрібно якнайбільше часу проводити на відкритих ділянках поза містами, поблизу річок, озер, у лісі, горах чи інших куточках природи. Про важливість цього фактора свідчить і те, що американське космічне агентство НАСА для забезпечення нормальної життєдіяльності астронавтів використовує спеціальні генератори хвиль Шумана. Наявність магнітного поля на Землі людина не відчуває, бо в неї немає для цього спеціального органу. Але без його існування життя на Землі в її сучасному вигляді неможливе. Таким чином, стрімка і значуща перебудова магнітного поля Землі не може проходити для нас безслідно.
Космічні фактори. Останніми десятиліттями важливе значення приділяється впливу космічних факторів на організми, популяції та екосистеми. Безумовно, одним із найважливіших космічних факторів є електромагнітне та корпускулярне випромінювання Сонця. Ще у 1915 році визначний вітчизняний учений XX століття А. Л. Чижев- ський сформулював положення про періодичний вплив Сонця на біосферу Землі, яке знаходить і надалі своє підтвердження. Учені відкрили цикли періодичної сонячної активності (корпускулярна радіація) у дуже широких інтервалах: від декількох хвилин і менше до десятків і сотень років. Із кількістю активних плям на Сонці пов’язані такі біологічні процеси, як врожайність сільськогосподарських культур, захворюваність населення, кількість спалахів масового розмноження різних організмів тощо.
У 1960-ті роки, коли штучний супутник облетів навколо Сонця, зареєстровано секторну природу міжпланетного магнітного поля (рис. 3.17). Послідовна зміна чотирьох секторів ММП, викликана сонячним вітром і обертанням Сонця навколо своєї осі, відбувається протягом 27 діб. Таким чином, приблизно через кожен тиждень відбувається зміна знаків ММП з «+» на «—» або навпаки. Добре відомо, що магнітне поле міжпланетного простору впливає на виникнення та протікання магнітних бур на Землі та на формування погоди.
Дослідження впливу змін міжпланетного секторного магнітного поля на стан біологічних і фізико-хімічних систем проводяться в Україні ученими Таврійського національного університету разом із працівниками Кримської астрофізичної обсерваторії. Встановлено, що разом зі зміною сектора МПП в організмах різних видів тварин, рослин, мікробів відбуваються істотні зміни: прискорюються або уповільнюються зростання та розвиток, змінюється активність багатьох ферментів тощо. Вивчення дії космічних чинників на організми, популяції, угруповання та біогеоценози — один із напрямів сучасної космічної екології, який успішно розвивається за кордоном і в Україні.
Рис. 3.17. Секторна структура міжпланетного магнітного поля в площині сонячного екватора; стрілками показано напрям силових ліній:можна бачити чотири сектори з постійним напрямом магнітного поля; межі між секторами проведені у вигляді спіралей Архімеда; надані дати та час переходу Землі у черговий сектор (Мизун, Мизун, 1984)
ВИКОРИСТАНА І РЕКОМЕНДОВАНА ЛІТЕРАТУРА
Білявський Г. О., Бутченко Л. І. Основи екології. — К.: Лібра, 2004. — 367 с.
Вальтер Г. Общая геоботаника. — М.: Мир, 1979.
Гиляров А. М. Популяционная экология. — М.: Изд-во МГУ, 1990. — 192 с.
Голубець М. А. Екосистемологія. — Львів: Поллі, 2000. — 316 с.
Голубець М. А. Середовищезнавство (інвайронментологія). — Львів: Манускрипт, 2010. — 176 с.
Джигирей В. С. Екологія та охорона навколишнього природного середовища. — К.: Знання, 2002. — 203 с.
Дідух Я. П., Ромащенко К. Ю. Теорія еконіші: вимір широти та перекриття // Укр. ботан. журн. — 2001. — Т. 58, № 5. — С. 529— 542.
Дідух Я. П. Етюди фітоекології. — К.: Арістей, 2008. — 264 с.
Космическая экология / В. Г. Сидякин, Н. А. Темурьянц, В. Б. Макеев, Б. М. Владимирский. — К.: Наукова думка, 1985. — 176 с.
Кучерявий В. П. Урбоекологія. — Львів: Світоч, 1999. — 359 с.
Кучерявий В. П. Загальна екологія. — Львів: Світ, 2010. — 520 с.
Словник найуживаніших термінів з екології, біотехнології і біоенергетики / М. Д. Мельничук, В. А. Гайченко, І. П. Григорюк та ін. — К.: Вид. НУБіП України, 2009. — 293 с.
Мизун Ю. Г., Мизун П. Г. Космос и здоровье. — М.: Знание, 1984. — 144 с.
Мусієнко М. М, Серебряков В. В., Брайон О. В. Екологія. Охорона природи: словник-довідник. — К.: Знання, 2002. — 551 с.
Пианка Э. Эволюционная экология. — М.: Мир, 1981. — 399 с.
Уиттекер Р. Сообщества и экосистемы. — М.: Прогресс, 1980. — 327 с.
Христофорова Н. К. Основы экологии. — Владивосток: Даль- наука, 1999. — 347 с.
Чернова Н. М., Былова А. М. Общая экология. — М.: Изд-во МГПУ, 1999. — 494 с.
Чижевский А. Л. Земное эхо солнечных бурь. — М.: Мысль, 1976. — 367 с.
Шилов И. А. Экология. —М.: Высшая школа, 2001. — 512 с.
Elton C. S. Animal Ecology. — London: Sidgwick and Jackson, 1927. — 209 p.
Grinnell J. The Niche Relationships of the California Transher. — Auk, 1917. — 21. — P. 364—382.
Dice L. R. Natural Communities. — Univ. Michigan Press, Ann Arbor., 1952. — 547 p.
Clarke G. L. Elements of Ecology. — New York: Hafner, 1954. — 560 p.
Hutchinson G. E. A Treatise on Limnology. Geography, Physics and Chemistry. — New York: Wiley, 1957. — Vol. 1. — 1015 p.
Hutchinson G. E. The Ecological Theater and the Evolutionary Play. — Yale Univ. Press, New Haven, Conn., 1965. — 139 p.
MaCarthur R. H. On the Relative Abundance of Bird Species. — Proc. Nat. Acad. Sci. 1957. — 43. — P. 293—295.
Odum E. P. Fundamentals of Ecology / 2nd ed. — Philadelphia: Saunders, 1959. — 564 p.
Pianka E. R. The Structure of Lizard Communities // Ann. Rev. Ecol., Syst. — 1973. — № 4. — P. 53—74.
ПИТАННЯ
ДЛЯ САМОСТІЙНОГО КОНТРОЛЮ ЗНАНЬ
1. Як зрозуміти поняття «оселище»?
2. Яка кількість екологічних чинників вам відома?
3. Чим відрізняються ресурси і умови?
4. Що таке адаптація?
5. Які екологічні чинники називають подразниками, обмежувачами, модифікаторами та сигналами?
6. Що таке «оптимум екологічного чинника»?
7. Які основні закономірності впливу екологічних чинників вам відомі?
8. Яку назву має явище порушення оптимуму відносно будь- якого чинника?
9. Як зрозуміти неспівпадання екологічних спектрів окремих видів?
10. Сформулюйте правило обмежуючих чинників.
11. Хто вперше запропонував термін «екологічна ніша»?
12. Чи можуть співіснувати види з однаковою екологічною нішею?
13. Дайте інтегральну характеристику екологічної ніші за Хатчинсоном.
14. Які відмінності існують між поняттями «фундаментальна ніша» та «реалізована ніша»?
15. Яким чином визначаються параметри екологічної ніші?
16. Який тип конкуренції розширює екологічну нішу, а який — звужує?
17. Сформулюйте закон максимуму щільності упаковки видів біоценозу при даних умовах середовища.
18. Чи залежить поведінка видів від характеру їх реалізованої екологічної ніші?
19. Які типи пристосування живих організмів до зовнішніх чинників існують?
20. Які закономірності описує правило оптимуму?
21. Що таке «екологічна валентність»?
22. Що таке екологічний оптимум і екологічний песимум?
23. Що означає термін «контамінація»?
24. Який вплив здійснюють модифікуючі чинники?
25. Що таке «лімітуючі чинники»?
26. У чому полягає принцип гомеостазу на рівні організму?
27. На які групи за принциповим екологічним значенням можна поділити адаптивні механізми?
28. Що відображають екологічні класифікації?
29. Хто вперше описав явище удаваної смерті й у чому воно полягає?
30. Яких властивостей набувають організми у стані анабіозу?
31. Чи є відмінності між гіпобіозом і криптобіозом?
32. Що визначає коефіцієнт Q10?
33. Чим відрізняються ендотермія та ектотермія?
34. Для яких організмів характерна гетеротермія?
35. Яке значення має сума ефективних температур?
36. Які механізми регуляції температури тіла притаманні пой- кілотермним тваринам?
37. Дайте визначення поняття базального метаболізму.
38. Чим відрізняється хімічна терморегуляція від фізичної?
39. Сформулюйте правило Бергмана та наведіть приклади його дії.
40. У чому полягає правило Аллена?
41. Які існують принципи екологічного поділу тварин за відношенням до світла?
42. Що таке «фотоперіодична регуляція»?
43. Які механізми лежать в основі циркадіанних (циркадних) ритмів?
44. Сформулюйте правило Ашоффа.
45. Які цикли називають циркануальними (цирканними)?
46. У чому полягає відмінність між циркануальними та сезонними ритмами?
47. На які екологічні зони поділяють Світовий океан?
48. Розшифруйте поняття «пойкілоосмотичні» та «гомойосмотичні» види тварин.
49. Які існують способи орієнтації тварин у водному середовищі?
50. Що таке анемохорія?
51. Які ґрунтові горизонти вам відомі? Надайте їх характеристику.
52. На які основні групи поділяють тварин — мешканців ґрунту?
53. Чим відрізняються «погода» і «клімат місцевості»?
54. У чому полягає важливість магнітного поля Землі для живих організмів?
55. Що таке хвилі Шумана і що відбувається з ними останнім часом?
56. Чому міській людині потрібно більше бувати на природі?
57. Що таке міжпланетне магнітне поле?
58. Як впливає міжпланетне магнітне поле на живі організми?
ТЕСТОВІ ЗАВДАННЯ
ДЛЯ САМОСТІЙНОГО КОНТРОЛЮ ЗНАНЬ
1. Основними механізмами адаптації на рівні організму є: а) онтогенетичні; b) популяційні; с) ценотичні.
2. Максимальні та мінімальні значення чинника, що може витримати організм — це: а) оптимальні точки; b) песимальні точки; с) критичні точки.
3. Нездатність переносити значні коливання тиску має назву:
а) стенотермність; b) стеногалінність; с) стенобатність; d) стено- топність.
4. Можливості існування організмів у середовищі обмежують ті чинники, які: а) найбільше віддалені від оптимуму; b) оптимальні;
c) найбільше наближені до оптимуму.
5. Термін «екологічна ніша» запропонував: а) Г. Гаузе; b) Ч. Ел- тон; с) Ж. Гріннел; d) Т. Шенер.
6. Фундаментальна ніша — той потенційно можливий гіпер- простір, який може зайняти вид за: а) відсутності конкуренції;
b) наявності конкуренції; с) зниженої конкуренції; d) підвищеної конкуренції.
7. За висловом Ю. Одума екологічна ніша — це: а) «адреса» виду; b) «професія» виду; с) «свідоцтво» виду.
8. Параметри еконіші визначають через показники: а) споживання ресурсів; b) наявності ресурсів; с) відсутності ресурсів;
d) конкуренції за ресурси.
9. За певними вимірами (факторами) еконіш високоспеціалі- зовані організми характеризуються межами толерантності, які є: а) вузькими; b) широкими; с) середніми; d) оптимальними.
10. У більшості випадків, чим більше видів у складі біоценозу, тим чисельність кожного із них: а) вища; b) нижча; с) не змінюється.
11. Адаптація за принципом толерантності — це шлях адаптації: а) пасивний; b) активний; с) кумулятивний; d) вибуховий.
12. Адаптація за принципом резистентності — це шлях адаптації:
a) пасивний; b) активний; с) кумулятивний; d) вибуховий.
13. У зоні оптимуму адаптивні механізми: а) посилені; b) послаблені; с) відключені; d) без змін.
14. Формулювання «Можливість існування даного виду в певному районі та ступінь його «процвітання» залежать від чинників, представлених у найменшій кількості» належить: а) Ю. Лібіху;
b) А. Тінеманну; с) Л. Раменському; d) В. Мізінгу.
15. Повна тимчасова зупинка життя отримала назву: а) гіпобіоз;
b) криптобіоз; с) анабіоз; d) гетеробіоз.
16. З ускладненням організації живих істот їх здатність бути активними за високих температур: а) знижується; b) підвищується;
c) не змінюється.
17. Терморегуляція внаслідок рефлекторного збільшення теплопродукції у відповідь на зниження температури середовища є: а) фізичною; b) біологічною; с) хімічною.
18. Тварини, для яких є характерними поведінкові способи регуляції теплообміну: а) пойкілотермні; b) гомойотермні; с) гетеро- термні; d) пойкілотермні і гомойотермні.
19. Частина світла, промені якої проникають у Світовий океан найглибше: а) довгохвильова; b) середньохвильова; с) короткохвильова; d) довгохвильова та середньохвильова.
20. Біологічні ритми з періодом близько 24 годин називаються: а) цирканними; b) циркадними; с) циркануальними.
21. Значна кількість мешканців морів і океанів відносно тиску є: а) еврибатними; b) полібатними; с) гетеробатними; d) стенобатними.
22. Багато видів водних тварин здатні за нестачі кисню впадати в неактивний стан, який називається: а) аноксибіозом; b) ан- гідробіозом; с) ангалінобіозом.
23. У гомойосмотичних тварин осмотичний тиск рідин тіла порівняно з навколишньою водою: а) вищий; b) нижчий; с) однаковий; d) не залежить від солоності води.
24. Середній розмір тварин, яких об’єднують під назвою мезофауна, складає: а) 0,1—3,0 мм; b) 2—20 мм; с) понад 20 мм.
25. Верхній, перегнійно-акумулятивний горизонт ґрунту, у якому накопичується і перетворюється органічна речовина, позначається літерою: а) А; b) В; с) С.
26. Положення про періодичний вплив Сонця на біосферу Землі сформулював: а) М. А. Голубець; b) В. І. Вернадський; с) А. Л. Чи- жевський.
27. Частота коливання магнітного поля Землі становить: а) 7 Гц; b) 7,8 Гц; с) наближається до 14 Гц.
ВІДПОВІДІ НА ТЕСТОВІ ЗАВДАННЯ
| № питання | Відповідь | № питання | Відповідь | № питання | Відповідь | № питання | Відповідь |
| 1 | a | 8 | a | 15 | с | 22 | a |
| 2 | с | 9 | a | 16 | a | 23 | d |
| 3 | с | 10 | b | 17 | с | 24 | a |
| 4 | a | 11 | a | 18 | d | 25 | a |
| 5 | с | 12 | b | 19 | с | 26 | с |
| 6 | a | 13 | с | 20 | b | 27 | с |
| 7 | b | 14 | a | 21 | d |
Еще по теме Найважливіші абіотичні фактори та адаптації до них:
- Абіотичні та біотичні фактори
- Загальні принципи адаптації
- Международные отношения на рубеже Средневековья и Нового времени. Место в них России
- Можливості адаптації організмів до токсикантів
- Інституалізація процесу адаптації українського законодавства до права Європейського Союзу
- 3. Стан та перспективи адаптації законодавства України до законодавства ЄС
- Екологічні фактори та їх класифікація
- Фактори середовища та закономірності їх дії на живі організми
- Можливість бути повноцінним учасником адміністративно-правових відносин та вільно задовольняти у них законні інтереси забезпечується наділенням фізичних, юридичних осіб, суб'єктів публічної адміністрації - імовірних учасників таких відносин - адміністративними повноваженнями.
- Етіологія та фактори ризику
- Етіологія та фактори ризику